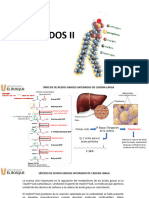
También podría gustarte
- 4.1 DislipoproteinemiasDocumento94 páginas4.1 DislipoproteinemiasDANIEL ALEJANDROAún no hay calificaciones
- Diapositivas - Química - LípidosDocumento143 páginasDiapositivas - Química - LípidosRaulRicardoCarhuapomaNicolasAún no hay calificaciones
- Lipidos IIIDocumento34 páginasLipidos IIIArriba GamersAún no hay calificaciones
- LÍPIDOSDocumento36 páginasLÍPIDOSgamarraexora034Aún no hay calificaciones
- Presentacion Final Degradación de LipidosDocumento33 páginasPresentacion Final Degradación de LipidosRoxy PeñalozaAún no hay calificaciones
- Quimica de Las MacromoleculasDocumento119 páginasQuimica de Las MacromoleculasCarlos Urgiles RollandAún no hay calificaciones
- CarbohidratosDocumento64 páginasCarbohidratosJuan camilo Gómez QAún no hay calificaciones
- Ebook Bioca 3ep 2023 LipidosDocumento18 páginasEbook Bioca 3ep 2023 LipidosC. Ibel Loza R.Aún no hay calificaciones
- VLDL, LDL, Idl, HDLDocumento23 páginasVLDL, LDL, Idl, HDLfarmacologicoAún no hay calificaciones
- BIOMOLECULASDocumento64 páginasBIOMOLECULASDuban Jose Galvis PradoAún no hay calificaciones
- Clase 9 - LIPIDOSDocumento46 páginasClase 9 - LIPIDOSpfloresangielisAún no hay calificaciones
- Lipidos II 2022Documento12 páginasLipidos II 2022Tatiana RodriguezAún no hay calificaciones
- BIOLOGÍA Resumen Cap. 3. LípidosDocumento5 páginasBIOLOGÍA Resumen Cap. 3. LípidosNadia N. SalattinoAún no hay calificaciones
- Archivo 9 Cartilla de Lipidos. Quimica BiologicaDocumento19 páginasArchivo 9 Cartilla de Lipidos. Quimica BiologicaRobin MaryAún no hay calificaciones
- Quimica Unidad 3 Carbohidratos CUP FCSH 2024Documento32 páginasQuimica Unidad 3 Carbohidratos CUP FCSH 2024Gerson Sanchez chuvéAún no hay calificaciones
- Sesión 03. Organización Molecular de La CélulaDocumento21 páginasSesión 03. Organización Molecular de La CélulaBelén SánchezAún no hay calificaciones
- BIOQUÍMICA p.1 PDFDocumento34 páginasBIOQUÍMICA p.1 PDFJohn Alejandro Mora PiernagordaAún no hay calificaciones
- Cap 9 Lípidos 2023Documento43 páginasCap 9 Lípidos 2023anaiflorcitaAún no hay calificaciones
- Lipidos Unidad 5Documento35 páginasLipidos Unidad 5Santiago RomeroAún no hay calificaciones
- 4 LípidosDocumento8 páginas4 LípidosCarmen Ochoa ValdésAún no hay calificaciones
- Movilización de Triacilgliceridos en MonogastricosDocumento11 páginasMovilización de Triacilgliceridos en Monogastricos50 EagleAún no hay calificaciones
- Clase 1. CarbohidratosDocumento43 páginasClase 1. CarbohidratosHerverth N LpzAún no hay calificaciones
- Metabolismo de LipidosDocumento76 páginasMetabolismo de LipidosErick Callenova MaytaAún no hay calificaciones
- C 2 U 5Documento15 páginasC 2 U 5Aida Navarrete RodriguezAún no hay calificaciones
- 13 Lípidos para PDFDocumento28 páginas13 Lípidos para PDFRenè OnaAún no hay calificaciones
- Semana 11 - CON APUNTESDocumento41 páginasSemana 11 - CON APUNTESLaura Diana Quispe aronesAún no hay calificaciones
- Biomoleculaslipidos Enf 2022Documento19 páginasBiomoleculaslipidos Enf 2022Melissa PintadoAún no hay calificaciones
- Lipidos Unidad IVDocumento19 páginasLipidos Unidad IVYASSIRA TAJA ROMEROAún no hay calificaciones
- LIPIDOSDocumento25 páginasLIPIDOSMinerva GilAún no hay calificaciones
- Lipidos 5Documento48 páginasLipidos 5kremenchutzkyAún no hay calificaciones
- Metabolismo Lípidos PDFDocumento42 páginasMetabolismo Lípidos PDFjose carlos jimenez huashuayoAún no hay calificaciones
- FisiopatologíasDocumento63 páginasFisiopatologíasDenis NavarroAún no hay calificaciones
- TEMA 6 Lipidos 1º Farma PB1 2019-2020 PARTE 1Documento19 páginasTEMA 6 Lipidos 1º Farma PB1 2019-2020 PARTE 1HelenaAún no hay calificaciones
- Presentacion LipidosDocumento12 páginasPresentacion LipidosGabrielAún no hay calificaciones
- TEMA No 7. LÍPIDOS PDFDocumento71 páginasTEMA No 7. LÍPIDOS PDFSantiago Loaiza PerezAún no hay calificaciones
- BIOMOLÉCULASDocumento18 páginasBIOMOLÉCULASNATALIA MERCEDES OLEA ACUÑAAún no hay calificaciones
- Clasificación de LipidosDocumento12 páginasClasificación de LipidosAndhy Ramirez MauriCioAún no hay calificaciones
- Repaso Parcial Todos Los Temas 1-7Documento13 páginasRepaso Parcial Todos Los Temas 1-7Pilar PazAún no hay calificaciones
- Lipidos RomyDocumento97 páginasLipidos Romyvelem1887Aún no hay calificaciones
- Bloque 3-BiomoleculasDocumento87 páginasBloque 3-BiomoleculasSalvador SoteloAún no hay calificaciones
- Biologia MolecularDocumento10 páginasBiologia MolecularPaula Valentina Sotelo PaezAún no hay calificaciones
- MoleculasDocumento10 páginasMoleculasapresaelizabethAún no hay calificaciones
- Unidad 1 Clasificación de Los Macronutrientes y MicronutrientesDocumento55 páginasUnidad 1 Clasificación de Los Macronutrientes y MicronutrientesDouglas SolorzanoAún no hay calificaciones
- PDF Lípidos IV EsfingoDocumento28 páginasPDF Lípidos IV EsfingoGian LuccaAún no hay calificaciones
- LIPIDOSDocumento10 páginasLIPIDOSlourdessbravooAún no hay calificaciones
- Manual Capacitacion LOPRAXINDocumento23 páginasManual Capacitacion LOPRAXINSonii Aguilar RiveraAún no hay calificaciones
- Semana 9-Sección 17Documento64 páginasSemana 9-Sección 17Ariana NicoleAún no hay calificaciones
- Lipidos - Apunte ConsolidadoDocumento15 páginasLipidos - Apunte Consolidadolotoflor2020Aún no hay calificaciones
- GLUCIDOSDocumento43 páginasGLUCIDOSanaliabelebrizuemaAún no hay calificaciones
- Mapa Conceptual Sindrome MetabolicoDocumento8 páginasMapa Conceptual Sindrome MetabolicoLeydi Portalanza VelardeAún no hay calificaciones
- Clase Sesion 7. Lipidos IDocumento40 páginasClase Sesion 7. Lipidos Ihermes alvarezAún no hay calificaciones
- Lipidos Saponificables e InsaponificablesDocumento36 páginasLipidos Saponificables e InsaponificablesBIANCA NATALIA ALVAREZ GUERREROAún no hay calificaciones
- LÍPIDOSDocumento57 páginasLÍPIDOSMilena AtaAún no hay calificaciones
- BiomoléculasDocumento8 páginasBiomoléculasIvoneVidal100% (1)
- Seminarios Quimica de Los AlimentosDocumento14 páginasSeminarios Quimica de Los Alimentosyw8nqff9vqAún no hay calificaciones
- Conf. Metab. de Lipoproteinas 15-16 Ile PDFDocumento32 páginasConf. Metab. de Lipoproteinas 15-16 Ile PDFWilson CoronadoAún no hay calificaciones
- Clase de Lípidos 2021Documento78 páginasClase de Lípidos 2021Ornella AlassiaAún no hay calificaciones
- Apuntes Bioquímica IIIDocumento33 páginasApuntes Bioquímica IIISTEPHANIE PAOLA VAZQUEZ ORTEGAAún no hay calificaciones
- Química de macrocomponentes de alimentosDe EverandQuímica de macrocomponentes de alimentosAún no hay calificaciones
- Seminario 10Documento42 páginasSeminario 10Aracely Escarleth Aguirre de la CruzAún no hay calificaciones
- Randy 3 FICHA DE PR CTICA 2 DEMOSTRACI N DE LAS LEYES DE MENDELDocumento6 páginasRandy 3 FICHA DE PR CTICA 2 DEMOSTRACI N DE LAS LEYES DE MENDELAracely Escarleth Aguirre de la CruzAún no hay calificaciones
- Semana 9-Sección 17Documento49 páginasSemana 9-Sección 17Aracely Escarleth Aguirre de la CruzAún no hay calificaciones
- Seminario 10Documento42 páginasSeminario 10Aracely Escarleth Aguirre de la CruzAún no hay calificaciones
- Semana 10Documento50 páginasSemana 10Aracely Escarleth Aguirre de la CruzAún no hay calificaciones
- Práctica 3 - Demostración Indirecta de Las Leyes de MendelDocumento5 páginasPráctica 3 - Demostración Indirecta de Las Leyes de MendelAracely Escarleth Aguirre de la CruzAún no hay calificaciones
- Seminario 2 GenéticaDocumento18 páginasSeminario 2 GenéticaAracely Escarleth Aguirre de la CruzAún no hay calificaciones
- Práctica 3 - Demostración Indirecta de Las Leyes de MendelDocumento5 páginasPráctica 3 - Demostración Indirecta de Las Leyes de MendelAracely Escarleth Aguirre de la CruzAún no hay calificaciones
- Seminario 2 GenéticaDocumento18 páginasSeminario 2 GenéticaAracely Escarleth Aguirre de la CruzAún no hay calificaciones
- Práctica 3 - Demostración Indirecta de Las Leyes de MendelDocumento5 páginasPráctica 3 - Demostración Indirecta de Las Leyes de MendelAracely Escarleth Aguirre de la CruzAún no hay calificaciones
- Práctica 3 - Demostración Indirecta de Las Leyes de MendelDocumento5 páginasPráctica 3 - Demostración Indirecta de Las Leyes de MendelAracely Escarleth Aguirre de la CruzAún no hay calificaciones
- Principales Criterios para La Aceptabilidad Ética de Los Estudios de Exposición Con Seres Humanos en El Contexto de La COVID-19Documento1 páginaPrincipales Criterios para La Aceptabilidad Ética de Los Estudios de Exposición Con Seres Humanos en El Contexto de La COVID-19Aracely Escarleth Aguirre de la CruzAún no hay calificaciones
- Randy 3 FICHA DE PR CTICA 2 DEMOSTRACI N DE LAS LEYES DE MENDELDocumento6 páginasRandy 3 FICHA DE PR CTICA 2 DEMOSTRACI N DE LAS LEYES DE MENDELAracely Escarleth Aguirre de la CruzAún no hay calificaciones
- 15 Participacion ComunitariaDocumento12 páginas15 Participacion ComunitariaAracely Escarleth Aguirre de la CruzAún no hay calificaciones
- 1 - 4963036286228103577 (1) - 28-29Documento2 páginas1 - 4963036286228103577 (1) - 28-29Aracely Escarleth Aguirre de la CruzAún no hay calificaciones
- Clase N1 Prevención Bioseguridad - Bacterias GeneralidadesDocumento76 páginasClase N1 Prevención Bioseguridad - Bacterias GeneralidadesAracely Escarleth Aguirre de la CruzAún no hay calificaciones
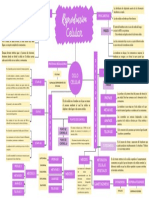
- Reproducción CelularDocumento47 páginasReproducción CelularFedra Stefany SalvadorAún no hay calificaciones
- Peach Wedding Plans Mind MapDocumento1 páginaPeach Wedding Plans Mind MapAracely Escarleth Aguirre de la CruzAún no hay calificaciones
- 1 - 4963036286228103577 (1) - 35-43Documento9 páginas1 - 4963036286228103577 (1) - 35-43Aracely Escarleth Aguirre de la CruzAún no hay calificaciones
- Peach Wedding Plans Mind MapDocumento1 páginaPeach Wedding Plans Mind MapAracely Escarleth Aguirre de la CruzAún no hay calificaciones
- Principales Criterios para La Aceptabilidad Ética de Los Estudios de Exposición Con Seres Humanos en El Contexto de La COVID-19. PAHO 2020Documento18 páginasPrincipales Criterios para La Aceptabilidad Ética de Los Estudios de Exposición Con Seres Humanos en El Contexto de La COVID-19. PAHO 2020Aracely Escarleth Aguirre de la CruzAún no hay calificaciones
- Microbiología e Inmunología-Bioseguridad-Semana 1-16Documento51 páginasMicrobiología e Inmunología-Bioseguridad-Semana 1-16Aracely Escarleth Aguirre de la CruzAún no hay calificaciones
- Sílabo Parasitología 2021 IIDocumento9 páginasSílabo Parasitología 2021 IIAracely Escarleth Aguirre de la CruzAún no hay calificaciones
- Sílabo Parasitología 2021 IIDocumento9 páginasSílabo Parasitología 2021 IIAracely Escarleth Aguirre de la CruzAún no hay calificaciones
- Microbiología e Inmunología-Bioseguridad-Semana 1-16Documento51 páginasMicrobiología e Inmunología-Bioseguridad-Semana 1-16Aracely Escarleth Aguirre de la CruzAún no hay calificaciones
- Clase N1 Prevención Bioseguridad - Bacterias GeneralidadesDocumento76 páginasClase N1 Prevención Bioseguridad - Bacterias GeneralidadesAracely Escarleth Aguirre de la CruzAún no hay calificaciones
- P2parasitos IntestinalesDocumento35 páginasP2parasitos IntestinalesAracely Escarleth Aguirre de la CruzAún no hay calificaciones
- P2parasitos IntestinalesDocumento35 páginasP2parasitos IntestinalesAracely Escarleth Aguirre de la CruzAún no hay calificaciones
- Semana 1 Morfo SistemasDocumento113 páginasSemana 1 Morfo SistemasVanessaAún no hay calificaciones
- Fenilalanina PDFDocumento12 páginasFenilalanina PDFEsteban Villegas ChávezAún no hay calificaciones
- Las VitaminasDocumento12 páginasLas VitaminasRomy C. FloresAún no hay calificaciones
- Práctica No.5 Inducción EnzimáticaDocumento10 páginasPráctica No.5 Inducción EnzimáticaAlis BretAAún no hay calificaciones
- Patogenia, Cribado y Diagnóstico de La Hipoglucemia Neonatal - UpToDateDocumento13 páginasPatogenia, Cribado y Diagnóstico de La Hipoglucemia Neonatal - UpToDatentytapAún no hay calificaciones
- DIETASCETOGENICASDocumento26 páginasDIETASCETOGENICASluz celeste herrera lopezAún no hay calificaciones
- Estres OxidativoDocumento9 páginasEstres Oxidativoricardo vallejosAún no hay calificaciones
- Drenaje y HomotoxicologíaDocumento9 páginasDrenaje y HomotoxicologíaRafa Rodriguez50% (2)
- S7 - L1. Biosíntesis de Ácidos Grasos y Eicosanoides PDFDocumento16 páginasS7 - L1. Biosíntesis de Ácidos Grasos y Eicosanoides PDFGaby Lisbeth LlhAún no hay calificaciones
- Guía para El Estudiante Fotosisnteiss y Respiración CelularDocumento5 páginasGuía para El Estudiante Fotosisnteiss y Respiración CelularValeria Alejandra Bravo RoblesAún no hay calificaciones
- Glosario de DeporteDocumento26 páginasGlosario de DeporteyelismarAún no hay calificaciones
- 4 LisosomasDocumento13 páginas4 LisosomasLety CatariAún no hay calificaciones
- TEMA 1. Introducción BiorreactoresDocumento5 páginasTEMA 1. Introducción BiorreactoresFátima Jerez ArroyoAún no hay calificaciones
- Beta Oxidación de Los Ácidos GrasosDocumento14 páginasBeta Oxidación de Los Ácidos GrasosISAAC MACARIO AVILA GARCIAAún no hay calificaciones
- Coenzimas REDOXDocumento30 páginasCoenzimas REDOXdescargalosAún no hay calificaciones
- Clase N8 (Semana 9 Respiracion Celular) Sesión 27 - 2020-2 PDFDocumento20 páginasClase N8 (Semana 9 Respiracion Celular) Sesión 27 - 2020-2 PDFRenzoAún no hay calificaciones
- Sensibilidad Del EtanolDocumento1 páginaSensibilidad Del EtanolPineda Lazaro AlexandraAún no hay calificaciones
- 5 Alimentos Que Destruyen Tu MetabolismoDocumento5 páginas5 Alimentos Que Destruyen Tu MetabolismoBRAULIO MATEO RAUDA RUIZAún no hay calificaciones
- 3 Clase Metabolismo MicrobianoDocumento30 páginas3 Clase Metabolismo MicrobianoOCTAVIOAún no hay calificaciones
- Fases de La FotosintesisDocumento1 páginaFases de La FotosintesisBolívar S. GutiérrezAún no hay calificaciones
- GonzalezGarcia Martin M14S3AI5Documento5 páginasGonzalezGarcia Martin M14S3AI5Lalo GonzálezAún no hay calificaciones
- Informe de Vitamina CDocumento10 páginasInforme de Vitamina CKaren SoledadAún no hay calificaciones
- Taller de Metabolismo de CarbohidratosDocumento8 páginasTaller de Metabolismo de CarbohidratosMARIA PAOLA GONZALEZ GONZALEZAún no hay calificaciones
- Equilibrio y BioenergeticaDocumento91 páginasEquilibrio y BioenergeticaMARIA CAMILA MARIN NIETOAún no hay calificaciones
- Metabolismo Vegetal - Fisiología VegetalDocumento9 páginasMetabolismo Vegetal - Fisiología VegetalCristoferVillarrealSantillanAún no hay calificaciones
- Taller #2 Esteban Diaz Fisiologia de La ActividadDocumento12 páginasTaller #2 Esteban Diaz Fisiologia de La ActividadEsteban alejandro Díaz bolañosAún no hay calificaciones
- Respiracion Celular - Obtencion de EnergiaDocumento23 páginasRespiracion Celular - Obtencion de EnergiaAbner HernandezAún no hay calificaciones
- InfografíaDocumento2 páginasInfografíamelissa giraldo100% (1)
- Caso Clinico 1 - Sala 1Documento2 páginasCaso Clinico 1 - Sala 1HxztoryAún no hay calificaciones
- El Thermogen Té o También Llamado TeatinoDocumento8 páginasEl Thermogen Té o También Llamado TeatinoWerbes HelenAún no hay calificaciones
- Etapas Del Anabolismo y CatabolismoDocumento8 páginasEtapas Del Anabolismo y CatabolismoAxel MelchorAún no hay calificaciones