También podría gustarte
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012De EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012Aún no hay calificaciones
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- Cadena Respiratoria IDocumento39 páginasCadena Respiratoria IZoila ParedesAún no hay calificaciones
- Parte de ExposiciónDocumento6 páginasParte de ExposiciónBaruch HernandezAún no hay calificaciones
- Cadena de Trasporte y Fosforilacion KKDocumento32 páginasCadena de Trasporte y Fosforilacion KKUriel AlvaradoAún no hay calificaciones
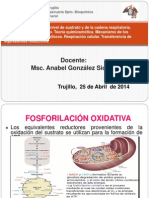
- Docente: Msc. Anabel González Siccha: Trujillo, 25 de Abril de 2014Documento30 páginasDocente: Msc. Anabel González Siccha: Trujillo, 25 de Abril de 2014Shantall SaquicorayAún no hay calificaciones
- Bioquimica Practica - Cadena RespiratoriaDocumento3 páginasBioquimica Practica - Cadena Respiratoriaingcivilunt1Aún no hay calificaciones
- Cadena Respiratoria IDocumento39 páginasCadena Respiratoria Irosse0% (1)
- Clase 17Documento5 páginasClase 17Pedro Esteban Ovalle AburtoAún no hay calificaciones
- Definición de Fosforilación OxidativaDocumento10 páginasDefinición de Fosforilación OxidativaDiomary ReyesAún no hay calificaciones
- 3-2022 Cte y Fosforilacion OxidativaDocumento56 páginas3-2022 Cte y Fosforilacion OxidativaAbisai FuentesAún no hay calificaciones
- Metabolismo Bacteriano GuiaDocumento7 páginasMetabolismo Bacteriano GuiaMay VonAún no hay calificaciones
- Discusion 4 Bioca IDocumento22 páginasDiscusion 4 Bioca Iestradaf6Aún no hay calificaciones
- Cadena Respiratoria OriginalDocumento31 páginasCadena Respiratoria OriginalTerry (Dino)Aún no hay calificaciones
- Bioquimic Fosforilacion OxidativaDocumento6 páginasBioquimic Fosforilacion OxidativapatriciaAún no hay calificaciones
- Resumen Completo de La Cadena Transportadora de Electrones y Fosforilación OxidativaDocumento3 páginasResumen Completo de La Cadena Transportadora de Electrones y Fosforilación OxidativaCarolina DuranAún no hay calificaciones
- Eq.3 Fosforilacion Oxidativa 2Documento24 páginasEq.3 Fosforilacion Oxidativa 2rubydonaldo0% (1)
- Cadena Respiración y Fosforilación Oxidativa Semana 8 CAE II 2022Documento14 páginasCadena Respiración y Fosforilación Oxidativa Semana 8 CAE II 2022vimoniAún no hay calificaciones
- La Cadena Respiratoria y Fosforilación OxidativaDocumento6 páginasLa Cadena Respiratoria y Fosforilación OxidativaCarlos VásquezAún no hay calificaciones
- Fosforilación OxidativaDocumento6 páginasFosforilación OxidativaLuis Manuel Arnaiz RodaAún no hay calificaciones
- Cadena Respiratoria y Fosforilación OxidativaDocumento6 páginasCadena Respiratoria y Fosforilación OxidativahenryAún no hay calificaciones
- Respiracion Celular 2Documento8 páginasRespiracion Celular 2Marlin TavarezAún no hay calificaciones
- Mecanismo Quimio-Osmótico de La Producción de AtpDocumento18 páginasMecanismo Quimio-Osmótico de La Producción de AtpKathe TafurAún no hay calificaciones
- La f1 f0 Atp SintasaDocumento8 páginasLa f1 f0 Atp SintasaAlexa MacielAún no hay calificaciones
- Discusion 4 Bioca IDocumento21 páginasDiscusion 4 Bioca IElena Donado50% (4)
- Respuestas Taller 4Documento3 páginasRespuestas Taller 4Mafer AraujoAún no hay calificaciones
- MitocondriasDocumento6 páginasMitocondriasDianaBGAún no hay calificaciones
- Cadena Respiratoria y Fosforilación OxidativaDocumento19 páginasCadena Respiratoria y Fosforilación OxidativaGreg Lopez100% (1)
- T-10 Teoria QuimiosmóticaDocumento30 páginasT-10 Teoria QuimiosmóticaUlises Benjamin Narro Altamirano0% (1)
- Fosforilacion OxidativaDocumento5 páginasFosforilacion Oxidativaaoihana18100% (1)
- Unidad Ii. Cadena Respiratoria y Fosforilacion Oxidativa PDFDocumento34 páginasUnidad Ii. Cadena Respiratoria y Fosforilacion Oxidativa PDFMafer AraujoAún no hay calificaciones
- Fosforilación OxidativaDocumento8 páginasFosforilación OxidativaLourdes PerdomoAún no hay calificaciones
- SESION 04. Bioenergética OkDocumento44 páginasSESION 04. Bioenergética Okprettyboy2898Aún no hay calificaciones
- FOSFORILACION OXIDATIVA (Recuperado Automáticamente)Documento4 páginasFOSFORILACION OXIDATIVA (Recuperado Automáticamente)Gutierrez Armenta Gabriela LizbethAún no hay calificaciones
- Bioca L Bloque 4 Discu 3Documento6 páginasBioca L Bloque 4 Discu 3TATIANAAún no hay calificaciones
- ApuntesDocumento20 páginasApuntesLucasAún no hay calificaciones
- Modelos para Explicar La Fosforilacion Oxidativa CompletoDocumento10 páginasModelos para Explicar La Fosforilacion Oxidativa CompletoCaArloss Adriian MarrtiinezzAún no hay calificaciones
- Guia KrebsDocumento9 páginasGuia KrebsVodkanicos OfficialAún no hay calificaciones
- Fosforilación Oxidativa - ResumenDocumento10 páginasFosforilación Oxidativa - ResumenKevinDelaCruzAún no hay calificaciones
- Cadena Transportadora de Electrones y Fosforilacion OxidativaDocumento29 páginasCadena Transportadora de Electrones y Fosforilacion OxidativaLoreto Carrasco AlarconAún no hay calificaciones
- 3 RESUMEN - Fosforilación OxidativaDocumento9 páginas3 RESUMEN - Fosforilación OxidativaMauricio DelgadilloAún no hay calificaciones
- Cadena Respiratoria, Fosforilacion Oxidativa Cap. 8 FerrierDocumento5 páginasCadena Respiratoria, Fosforilacion Oxidativa Cap. 8 FerrierEvelyn Guadalupe González BravoAún no hay calificaciones
- Fosforilación OxidativaDocumento12 páginasFosforilación OxidativaJudit BoixAún no hay calificaciones
- Cadena RespiratoriaDocumento27 páginasCadena RespiratoriaRicardo FonsecaAún no hay calificaciones
- Inhibidores Enzimaticos en La Cadena RespiratoriaDocumento5 páginasInhibidores Enzimaticos en La Cadena RespiratoriaFATIMA GERALDINE PIZAN OCUPAAún no hay calificaciones
- Bioquimica FosforilaciónDocumento35 páginasBioquimica FosforilaciónJaime Campos HernándezAún no hay calificaciones
- Tema 3Documento15 páginasTema 3juanmideluxeAún no hay calificaciones
- 7.1 Fosforilacion Oxidativa y FotosforilacionDocumento21 páginas7.1 Fosforilacion Oxidativa y FotosforilacionDiana Diana ValenzuelaAún no hay calificaciones
- Fosforilación OxidativaDocumento7 páginasFosforilación Oxidativadarco16Aún no hay calificaciones
- Fosforilación OxidativaDocumento20 páginasFosforilación OxidativaValeri Vasquez Cucho0% (1)
- Ubicación de La ClorofilaDocumento3 páginasUbicación de La ClorofilaTahic PAún no hay calificaciones
- Bioenergetica Fotosintesis 2019Documento51 páginasBioenergetica Fotosintesis 2019Antonela SorucoAún no hay calificaciones
- Fosforilación OxidativaDocumento2 páginasFosforilación OxidativaNEIVERAún no hay calificaciones
- Caso Clinico 2Documento14 páginasCaso Clinico 2ANNIE NICOLL ZAVALETA QUIROZAún no hay calificaciones
- Fosforilación OxidativaDocumento26 páginasFosforilación OxidativaXiomara Sanchez CahuanaAún no hay calificaciones
- Tema 8 y 9Documento25 páginasTema 8 y 9halley moonAún no hay calificaciones
- Ashley Salcedo 100478923 Bioquimica 2Documento5 páginasAshley Salcedo 100478923 Bioquimica 2Lenny MotaAún no hay calificaciones
- Fosforilacion Oxidativa Teoria QuimiosmoticaDocumento18 páginasFosforilacion Oxidativa Teoria QuimiosmoticaJose Arceo CendejasAún no hay calificaciones
- BioquímicaDocumento3 páginasBioquímicaKarold VanessaAún no hay calificaciones
- Introducción a la Biología: RESÚMENES UNIVERSITARIOSDe EverandIntroducción a la Biología: RESÚMENES UNIVERSITARIOSCalificación: 5 de 5 estrellas5/5 (1)
- 4to Parcial Clase de Introduccion A La ClinicaDocumento6 páginas4to Parcial Clase de Introduccion A La ClinicaNeris ColladoAún no hay calificaciones
- Genetica ManualDocumento24 páginasGenetica ManualSugeiry FriasAún no hay calificaciones
- Examenes de Fisiologia 2Documento10 páginasExamenes de Fisiologia 2Sugeiry FriasAún no hay calificaciones
- Guyton Regulación AcidobásicaDocumento18 páginasGuyton Regulación AcidobásicaSugeiry FriasAún no hay calificaciones
- Historia de La RadiologíaDocumento13 páginasHistoria de La RadiologíaSugeiry FriasAún no hay calificaciones
- GenéticaDocumento1 páginaGenéticaSugeiry FriasAún no hay calificaciones
- PUENTEDocumento15 páginasPUENTESugeiry FriasAún no hay calificaciones
- DIENCÉFALODocumento14 páginasDIENCÉFALOSugeiry FriasAún no hay calificaciones
- Resumen 3erppDocumento36 páginasResumen 3erppSugeiry FriasAún no hay calificaciones
- Función PolinómicaDocumento1 páginaFunción PolinómicaSugeiry FriasAún no hay calificaciones
- Anomalías CongénitasDocumento16 páginasAnomalías CongénitasSugeiry FriasAún no hay calificaciones
- BulboDocumento14 páginasBulboSugeiry FriasAún no hay calificaciones
- GineDocumento8 páginasGineSugeiry FriasAún no hay calificaciones
- Area Del Visual Terciara Area Sensitiva Primaria Area Motora PrimariaDocumento4 páginasArea Del Visual Terciara Area Sensitiva Primaria Area Motora PrimariaSugeiry FriasAún no hay calificaciones
- Area Del Visual Terciara Area Sensitiva Primaria Area Motora PrimariaDocumento4 páginasArea Del Visual Terciara Area Sensitiva Primaria Area Motora PrimariaSugeiry FriasAún no hay calificaciones
- Peritoneo y Cavidad PeritonealDocumento3 páginasPeritoneo y Cavidad PeritonealsharollAún no hay calificaciones
- UntitledDocumento12 páginasUntitledSugeiry FriasAún no hay calificaciones
- El Bazo y PancreaDocumento2 páginasEl Bazo y PancreaSugeiry FriasAún no hay calificaciones
- Sustancia Blanca: Cerebro Pesa 1,100 1,300 GramosDocumento4 páginasSustancia Blanca: Cerebro Pesa 1,100 1,300 GramosSugeiry FriasAún no hay calificaciones
- Hormona PlacentariaDocumento3 páginasHormona PlacentariaSugeiry FriasAún no hay calificaciones
- EsófagoDocumento3 páginasEsófagoSugeiry FriasAún no hay calificaciones
- Diagnostico Presuntivo y DefinitivoDocumento2 páginasDiagnostico Presuntivo y DefinitivoSugeiry FriasAún no hay calificaciones
- Irrigación Del PeritoneoDocumento3 páginasIrrigación Del PeritoneoSugeiry FriasAún no hay calificaciones
- Importancia Del PH en Nuestro OrganismoDocumento4 páginasImportancia Del PH en Nuestro Organismocecilia yamunaquesilvaAún no hay calificaciones
- Guia Once Funciones Oxigenadas 2 ParteDocumento24 páginasGuia Once Funciones Oxigenadas 2 Partesergio alarconAún no hay calificaciones
- CATALOGOBUNGARDDocumento32 páginasCATALOGOBUNGARDSander DuqueAún no hay calificaciones
- Composicion de Placa Microbiana de La Cavidad OralDocumento4 páginasComposicion de Placa Microbiana de La Cavidad OralMilena CoppianoAún no hay calificaciones
- Methanobrevibacter RuminantiumDocumento2 páginasMethanobrevibacter RuminantiumEstefaníaCeballosAún no hay calificaciones
- TALLER CorrosionDocumento5 páginasTALLER CorrosionshelsiAún no hay calificaciones
- Practica 2 Manejo de Residuos SolidosDocumento3 páginasPractica 2 Manejo de Residuos SolidosCAMILE ALEJANDRA CACERES SALASAún no hay calificaciones
- Control Union - Comparativa CERTIFICACIONESDocumento26 páginasControl Union - Comparativa CERTIFICACIONESJuan IgnacioAún no hay calificaciones
- Presentacion Bombilleria Dic 2022Documento5 páginasPresentacion Bombilleria Dic 2022Dar FallAún no hay calificaciones
- Anual Uni-Semana 03 QuímicaDocumento26 páginasAnual Uni-Semana 03 QuímicaMarco Antonio Palacios ReyesAún no hay calificaciones
- Tema 4 Limpieza, Desinfección y Esterilización Curso 2324Documento15 páginasTema 4 Limpieza, Desinfección y Esterilización Curso 2324martablet506Aún no hay calificaciones
- Metabolismo Secundario en PlantasDocumento1 páginaMetabolismo Secundario en Plantasvalentina leal100% (1)
- 4.1-Estado de OxidacionDocumento5 páginas4.1-Estado de OxidacionJeyson GalaAún no hay calificaciones
- Efecto FantasmaDocumento3 páginasEfecto Fantasmapirotecnia scmixAún no hay calificaciones
- Mechero Bunsen - Laboratorio de QuímicaDocumento18 páginasMechero Bunsen - Laboratorio de QuímicaDJPalanca 19Aún no hay calificaciones
- Tema 7 Tipos de ContaminacionDocumento5 páginasTema 7 Tipos de ContaminacionKaren Rosa Ramirez VillcaAún no hay calificaciones
- Jabon de Zanahoria Informe 1Documento21 páginasJabon de Zanahoria Informe 1Karla MostaceroAún no hay calificaciones
- ReportePractica Practica7 LopezTorresDocumento10 páginasReportePractica Practica7 LopezTorresJennifer Gabriela Lopez TorresAún no hay calificaciones
- Manual de Nomenclatura OrgánicaDocumento5 páginasManual de Nomenclatura OrgánicaGabriel CastañedaAún no hay calificaciones
- Tartrato de Antimonio PotasioDocumento8 páginasTartrato de Antimonio PotasioCristina Nicole VelázquezAún no hay calificaciones
- 1 - Ficha Teìcnica HALOTRON 1Documento1 página1 - Ficha Teìcnica HALOTRON 1Jonathan RamírezAún no hay calificaciones
- Naturelle S2 Hydraulic Fluid 32Documento3 páginasNaturelle S2 Hydraulic Fluid 32Edgar Leonel CortésAún no hay calificaciones
- Extracción de Aceite de Higuerilla - Descripción Del ProcesoDocumento44 páginasExtracción de Aceite de Higuerilla - Descripción Del ProcesodanielAún no hay calificaciones
- Bacteria y Su Clasificacion.Documento21 páginasBacteria y Su Clasificacion.Maria josefa Garcia BayonaAún no hay calificaciones
- Cuestionario Laboratorio 2Documento7 páginasCuestionario Laboratorio 2Noelia Alondra MartinezAún no hay calificaciones
- Procedimientos de Seguridad en Trabajos Críticos y de Alto RiesgoDocumento26 páginasProcedimientos de Seguridad en Trabajos Críticos y de Alto RiesgoDaniel Enrique Horna HernándezAún no hay calificaciones
- Taller Refuerzo Qui 2023Documento3 páginasTaller Refuerzo Qui 2023Luciana pAún no hay calificaciones
- Informe N°9-Rendimiento PorcentualDocumento6 páginasInforme N°9-Rendimiento PorcentualKOKE MCAún no hay calificaciones
- Tablas de WPC A Partir de Residuos de AserrínDocumento18 páginasTablas de WPC A Partir de Residuos de AserrínValeria Bedoya Jaramillo100% (1)
- Análisis SaraDocumento52 páginasAnálisis SaraMax Cedeño De León100% (1)