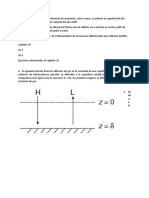
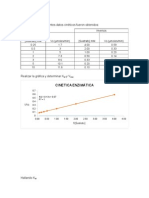
También podría gustarte
- EnzimasDocumento28 páginasEnzimasAna FilichAún no hay calificaciones
- Enzimas WordDocumento14 páginasEnzimas WordYaz ReyesAún no hay calificaciones
- Bloque 2 BioquímicaDocumento23 páginasBloque 2 BioquímicaSergio SeseteAún no hay calificaciones
- Rutas MetabolicasDocumento33 páginasRutas MetabolicasJamie Jayjay Martinez100% (2)
- EnzimasDocumento27 páginasEnzimasYAMILETH ACOSTA GUTIERREZAún no hay calificaciones
- EnzimasDocumento23 páginasEnzimasMaruby MontonerAún no hay calificaciones
- Practica No. 3. Actividad Enzimatica - AmilasaDocumento12 páginasPractica No. 3. Actividad Enzimatica - AmilasanicoleAún no hay calificaciones
- ENZIMASDocumento11 páginasENZIMASjuliaAún no hay calificaciones
- 10 EnzimasDocumento16 páginas10 EnzimasMiguel MancusoAún no hay calificaciones
- Primera Ley de La TermodinámicaDocumento34 páginasPrimera Ley de La TermodinámicaGedrik Denilson Kluivert Segura PineloAún no hay calificaciones
- TEMA 4bioqDocumento9 páginasTEMA 4bioqAlba Montes FernandezAún no hay calificaciones
- Enzimas 1Documento116 páginasEnzimas 1JM JessycaAún no hay calificaciones
- Cinética EnzimáticaDocumento9 páginasCinética EnzimáticaLourdes SoriaAún no hay calificaciones
- Sanmartin Enzimas 3a AgronomiaDocumento8 páginasSanmartin Enzimas 3a AgronomiaJOSUE TADEO SANMARTIN JUMBOAún no hay calificaciones
- EnzimasDocumento95 páginasEnzimas3F Rodríguez Alcocer NataliaAún no hay calificaciones
- Tema 6. Enzimas.Documento12 páginasTema 6. Enzimas.DCPAún no hay calificaciones
- Biocatalizadores y Referencias BibliograficasDocumento6 páginasBiocatalizadores y Referencias BibliograficasMaria Elizabeth M. Alberca0% (1)
- Bioquimica Ezimas y BioenergeticaDocumento20 páginasBioquimica Ezimas y Bioenergeticajulia campoAún no hay calificaciones
- C5 - Metabolismo CelularDocumento4 páginasC5 - Metabolismo CelularMarcos NovoAún no hay calificaciones
- 5 - EnzimasDocumento7 páginas5 - Enzimasnahiarabeluu2Aún no hay calificaciones
- EnzimasDocumento9 páginasEnzimasFernando Rodriguez ChavezAún no hay calificaciones
- Cap 7 - Enzimas - Mecanismo de AcciónDocumento12 páginasCap 7 - Enzimas - Mecanismo de AcciónCharlesAún no hay calificaciones
- Unidad 6.1 - Cinética Enzimática-PresentaciónDocumento74 páginasUnidad 6.1 - Cinética Enzimática-PresentaciónCamila EscobarAún no hay calificaciones
- Las enzimas catalizan reacciones químicasDocumento17 páginasLas enzimas catalizan reacciones químicasMartina MoralesAún no hay calificaciones
- Enzimas PDFDocumento8 páginasEnzimas PDFAnonymous KarinaAún no hay calificaciones
- EnzimasDocumento12 páginasEnzimasmakoki92Aún no hay calificaciones
- MoneycoachDocumento24 páginasMoneycoachackermamyusselAún no hay calificaciones
- Enzimas Marco TeoricoDocumento4 páginasEnzimas Marco TeoricoHenrry Maldonado SedanoAún no hay calificaciones
- Tema 3 - EnzimasDocumento3 páginasTema 3 - EnzimasMari Carmen ReinéAún no hay calificaciones
- Apuntes de EnzimasDocumento4 páginasApuntes de EnzimasYadira Ramirez ReyesAún no hay calificaciones
- EnzimasDocumento8 páginasEnzimasmetroid999Aún no hay calificaciones
- 1. EnzimasDocumento3 páginas1. EnzimasNatasha GabrielleAún no hay calificaciones
- Células y seres vivos como sistemas abiertos en equilibrio dinámicoDocumento5 páginasCélulas y seres vivos como sistemas abiertos en equilibrio dinámicoAna Valadés AlcarazAún no hay calificaciones
- Taller de BioquimicaDocumento8 páginasTaller de BioquimicaYeicid Carolina Pabon PerezAún no hay calificaciones
- Metabolismo CelularDocumento9 páginasMetabolismo CelularamandaAún no hay calificaciones
- MetabolismoDocumento16 páginasMetabolismoNatalia AssoAún no hay calificaciones
- Clasificación y mecanismos de las enzimasDocumento22 páginasClasificación y mecanismos de las enzimasFercho MariscalAún no hay calificaciones
- Aprendizaje 10Documento8 páginasAprendizaje 10ricardo moralesAún no hay calificaciones
- t2 enzimasDocumento40 páginast2 enzimashbetancor2005Aún no hay calificaciones
- Enzimas y Sutratos. FisiologíaDocumento16 páginasEnzimas y Sutratos. FisiologíaFaustino MarsoAún no hay calificaciones
- ENZIMASDocumento3 páginasENZIMAShola soy agustinaAún no hay calificaciones
- Guia 1 BX EnzimasDocumento12 páginasGuia 1 BX EnzimasYamari MartinezAún no hay calificaciones
- El Metabolismo y EnzimasDocumento42 páginasEl Metabolismo y EnzimasGuillermo ChavesAún no hay calificaciones
- ENZIMASDocumento11 páginasENZIMASamilcar torresAún no hay calificaciones
- Cuestionario Enzimas UPCDocumento6 páginasCuestionario Enzimas UPCLUZ DARY SERRANO SERRANOAún no hay calificaciones
- Enzimas Mecanismo de AcciónDocumento10 páginasEnzimas Mecanismo de AcciónNayeli LasluisaAún no hay calificaciones
- Revisión bibliográfica sobre enzimologíaDocumento57 páginasRevisión bibliográfica sobre enzimologíaNaranjoAlvarezCristianAún no hay calificaciones
- 1.8 Energetica de La VidaDocumento8 páginas1.8 Energetica de La VidaKevin MoralesAún no hay calificaciones
- Enzimas Agro1 2023Documento52 páginasEnzimas Agro1 2023202208134Aún no hay calificaciones
- Apuntes Clase 2 BiocatalisisDocumento3 páginasApuntes Clase 2 Biocatalisisyasmin zavaleta velezAún no hay calificaciones
- Cofactores MetálicosDocumento6 páginasCofactores MetálicosFiorella Alexandra0% (1)
- Taller de Repaso de EnzimasDocumento8 páginasTaller de Repaso de EnzimasDani CardonaAún no hay calificaciones
- EnzimasDocumento11 páginasEnzimasvanesaAún no hay calificaciones
- Ejercicio 1 EnzimologiaDocumento15 páginasEjercicio 1 EnzimologiaAndrea ChavarroAún no hay calificaciones
- Taller de Repaso de EnzimasDocumento8 páginasTaller de Repaso de EnzimasDani CardonaAún no hay calificaciones
- Enzimas: Actividad y factores que la afectanDocumento3 páginasEnzimas: Actividad y factores que la afectanSoy un Gordo ConchatumadreAún no hay calificaciones
- En ZimasDocumento5 páginasEn Zimasbryan gomez de la cruzAún no hay calificaciones
- Enzimas 2Documento19 páginasEnzimas 2Eduardo FeriaAún no hay calificaciones
- Resumen y Comentario Crítico Texto 1 - Lara Iglesias PDFDocumento2 páginasResumen y Comentario Crítico Texto 1 - Lara Iglesias PDFLara MaríaAún no hay calificaciones
- Regulación enzimática enDocumento4 páginasRegulación enzimática enLara MaríaAún no hay calificaciones
- Tema 8 - BioenergéticaDocumento11 páginasTema 8 - BioenergéticaLara MaríaAún no hay calificaciones
- Cinética EnzimáticaDocumento5 páginasCinética EnzimáticaLara MaríaAún no hay calificaciones
- Tema 6 - Función de ProteínasDocumento5 páginasTema 6 - Función de ProteínasLara MaríaAún no hay calificaciones
- Tema 7 - Membranas Biológicas y TransporteDocumento12 páginasTema 7 - Membranas Biológicas y TransporteLara MaríaAún no hay calificaciones
- Tema 4 - Nucleótidos y AminoácidosDocumento5 páginasTema 4 - Nucleótidos y AminoácidosLara MaríaAún no hay calificaciones
- Tema 5 - Enlace Peptídico y ProteínasDocumento5 páginasTema 5 - Enlace Peptídico y ProteínasLara MaríaAún no hay calificaciones
- Tema 3 - LípidosDocumento6 páginasTema 3 - LípidosLara MaríaAún no hay calificaciones
- Tema 1 - La Materia Viva y Los IsómerosDocumento3 páginasTema 1 - La Materia Viva y Los IsómerosLara MaríaAún no hay calificaciones
- Glúcidos y carbohidratos: clasificación y propiedadesDocumento8 páginasGlúcidos y carbohidratos: clasificación y propiedadesLara MaríaAún no hay calificaciones
- Planificación anual de química 3o medio 2020Documento9 páginasPlanificación anual de química 3o medio 2020Ana María Millán InzunzaAún no hay calificaciones
- Catalizadores Heterogéneos Utilizados para La Obtención de Biodiesel Exposicion.Documento3 páginasCatalizadores Heterogéneos Utilizados para La Obtención de Biodiesel Exposicion.andreaAún no hay calificaciones
- Xylene IsomerizationDocumento5 páginasXylene IsomerizationLuis Alberto Flores RiojaAún no hay calificaciones
- Taller Transferencia de MasaDocumento4 páginasTaller Transferencia de MasaDavidsantiago Murillo AvilaAún no hay calificaciones
- Equilibrio y Cinetica QuimicaDocumento22 páginasEquilibrio y Cinetica QuimicaMatías Valenzuela NachbauerAún no hay calificaciones
- Competencias: CapítuloDocumento67 páginasCompetencias: CapítuloYuly Condezo CabreraAún no hay calificaciones
- Perspectivas FuturasDocumento44 páginasPerspectivas FuturasGabriela OjedaAún no hay calificaciones
- Instrucciones Detalladas para Hacer Goma EspumaDocumento27 páginasInstrucciones Detalladas para Hacer Goma EspumaJohn FredyAún no hay calificaciones
- Jitorres - 1. Talleres Reciclo y Purga. SelectividadDocumento1 páginaJitorres - 1. Talleres Reciclo y Purga. SelectividadCamilo VanegasAún no hay calificaciones
- Formacion de CumarinasDocumento15 páginasFormacion de CumarinasEstephany Ortega0% (1)
- Sesión 05 Cinética QuímicaDocumento36 páginasSesión 05 Cinética QuímicaAdolfo GuerreroAún no hay calificaciones
- Diseño Base de Planta PG Natalia SanabriaDocumento36 páginasDiseño Base de Planta PG Natalia SanabriaNATALIA SANABRIA ALBAAún no hay calificaciones
- Efecto Del PH y Temperatura Sobre La Actividad Enzimatica de La Catalasa y PeroxidasaDocumento41 páginasEfecto Del PH y Temperatura Sobre La Actividad Enzimatica de La Catalasa y PeroxidasaGustavo100% (2)
- Trabajo 2Documento2 páginasTrabajo 2Lizeth Martinez RamirezAún no hay calificaciones
- PiaDocumento9 páginasPiamonyAún no hay calificaciones
- Semana 12 Enzimas IDocumento26 páginasSemana 12 Enzimas IJanneth TubonAún no hay calificaciones
- Energia de ActivacionDocumento2 páginasEnergia de ActivacionIris Azucena Fernández SalazarAún no hay calificaciones
- Los oligoelementos catalizadores: introducción a la medicina funcionalDocumento20 páginasLos oligoelementos catalizadores: introducción a la medicina funcionalfisijazz86% (7)
- Ejercicios de Cinetica EnzimaticaDocumento7 páginasEjercicios de Cinetica EnzimaticaJoseChesireAún no hay calificaciones
- Practico de Refinacion 3Documento10 páginasPractico de Refinacion 3marvinAún no hay calificaciones
- 3 Medio - Quimica - Zig Zag - EstudianteDocumento200 páginas3 Medio - Quimica - Zig Zag - EstudianteKarla Daniela Muñoz Martinez100% (1)
- Traduccion Del HipopotamoDocumento14 páginasTraduccion Del HipopotamoGerardo GomezAún no hay calificaciones
- Diseño Básico de Procesos: Etapas y ConsideracionesDocumento42 páginasDiseño Básico de Procesos: Etapas y ConsideracionesYelo Bans RaMenAún no hay calificaciones
- HIDROLISISDocumento12 páginasHIDROLISISYesenia Barrios Ccoyori100% (1)
- Catalizadores HZSM-5 para mejorar bio-aceite de pirólisisDocumento3 páginasCatalizadores HZSM-5 para mejorar bio-aceite de pirólisisAngelo ToledoAún no hay calificaciones
- Foss Tecator DigestorDocumento10 páginasFoss Tecator DigestorAnonymous kTtClaOLAAún no hay calificaciones
- Ejercicio 1 EnzimologiaDocumento15 páginasEjercicio 1 EnzimologiaAndrea ChavarroAún no hay calificaciones
- Soluciones Test Repaso Unidad 4Documento3 páginasSoluciones Test Repaso Unidad 4lauraAún no hay calificaciones
- Esta Di SticaDocumento11 páginasEsta Di Sticaedinsonduran100% (3)
- Anatomía & 100 estiramientos Esenciales (Color): Técnicas, beneficios, precauciones, consejos, tablas de series, dolenciasDe EverandAnatomía & 100 estiramientos Esenciales (Color): Técnicas, beneficios, precauciones, consejos, tablas de series, dolenciasCalificación: 4.5 de 5 estrellas4.5/5 (21)
- La revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaDe EverandLa revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaCalificación: 5 de 5 estrellas5/5 (200)
- La metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceDe EverandLa metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceCalificación: 5 de 5 estrellas5/5 (8)
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- Disciplina Mental: Técnicas infalibles para lograr todo lo que te propones y eliminar la pereza y la procrastinación de tu vida para siempreDe EverandDisciplina Mental: Técnicas infalibles para lograr todo lo que te propones y eliminar la pereza y la procrastinación de tu vida para siempreCalificación: 5 de 5 estrellas5/5 (3)
- Zensorialmente : Dejá que tu cuerpo sea tu cerebroDe EverandZensorialmente : Dejá que tu cuerpo sea tu cerebroAún no hay calificaciones
- Fisiopatología de las enfermedades cardiovascularesDe EverandFisiopatología de las enfermedades cardiovascularesCalificación: 5 de 5 estrellas5/5 (1)
- Desastres y emergencias. Prevención, mitigación y preparaciónDe EverandDesastres y emergencias. Prevención, mitigación y preparaciónCalificación: 4 de 5 estrellas4/5 (5)
- La invasión de la pseudociencia: Claves para orientarse en un mundo en donde casi todo es posibleDe EverandLa invasión de la pseudociencia: Claves para orientarse en un mundo en donde casi todo es posibleCalificación: 4.5 de 5 estrellas4.5/5 (15)
- Neurociencia para vencer la depresión: La esprial ascendenteDe EverandNeurociencia para vencer la depresión: La esprial ascendenteCalificación: 4.5 de 5 estrellas4.5/5 (10)
- Interpretación del ECG: Una Guía Práctica e Intuitiva para Aprender a Leer el ECG y Diagnosticar y Tratar ArritmiasDe EverandInterpretación del ECG: Una Guía Práctica e Intuitiva para Aprender a Leer el ECG y Diagnosticar y Tratar ArritmiasAún no hay calificaciones
- Reconstrucción de dientes endodonciados: Pautas de actuación clínicaDe EverandReconstrucción de dientes endodonciados: Pautas de actuación clínicaCalificación: 5 de 5 estrellas5/5 (4)
- El libro conciso de la punción seca: Manual del terapeuta para las aplicaciones en los puntos gatillo miofasciales (Color)De EverandEl libro conciso de la punción seca: Manual del terapeuta para las aplicaciones en los puntos gatillo miofasciales (Color)Calificación: 3 de 5 estrellas3/5 (2)
- Trucos para Leer la Mente de los Demás: Cómo Adivinar el Pensamiento de los Demás con Poco Esfuerzo. 2 Libros en 1 - Secretos de la Psicología Oscura, Cómo ser un Detector de MentirasDe EverandTrucos para Leer la Mente de los Demás: Cómo Adivinar el Pensamiento de los Demás con Poco Esfuerzo. 2 Libros en 1 - Secretos de la Psicología Oscura, Cómo ser un Detector de MentirasCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Borges y la memoria: De "Funes el memorioso" a la neurona de Jennifer AnistonDe EverandBorges y la memoria: De "Funes el memorioso" a la neurona de Jennifer AnistonCalificación: 5 de 5 estrellas5/5 (4)
- Cómo hacer aviones de papel y otros objetos voladoresDe EverandCómo hacer aviones de papel y otros objetos voladoresAún no hay calificaciones
- El péndulo de sanación: Péndulo hebreo. Investigación y sistematización de la técnicaDe EverandEl péndulo de sanación: Péndulo hebreo. Investigación y sistematización de la técnicaCalificación: 4.5 de 5 estrellas4.5/5 (27)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Prescripción de ejercico físico para la saludDe EverandPrescripción de ejercico físico para la saludCalificación: 5 de 5 estrellas5/5 (1)
- Notas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesDe EverandNotas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesAún no hay calificaciones
- Dieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasDe EverandDieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasCalificación: 4 de 5 estrellas4/5 (9)
- Teoría de juegos: Una introducción matemática a la toma de decisionesDe EverandTeoría de juegos: Una introducción matemática a la toma de decisionesCalificación: 4 de 5 estrellas4/5 (1)