También podría gustarte
- Demostracion de Cuerpos Cetonicos en La OrinaDocumento21 páginasDemostracion de Cuerpos Cetonicos en La OrinaYiselda LisetheAún no hay calificaciones
- DISLIPIDEMIASDocumento21 páginasDISLIPIDEMIASAlbina Pamo100% (1)
- Estatus ácido-base: Conceptos, desequilibrios e interpretaciónDe EverandEstatus ácido-base: Conceptos, desequilibrios e interpretaciónAún no hay calificaciones
- Metabolismo de Las LipoproteinasDocumento12 páginasMetabolismo de Las LipoproteinasMargot Lima Ccahuana100% (1)
- Practica N°8 Perfil Lipidico Del Suero 2020Documento47 páginasPractica N°8 Perfil Lipidico Del Suero 2020Fernando ZapanaAún no hay calificaciones
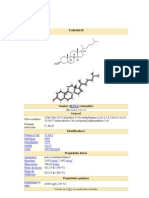
- ColesterolDocumento11 páginasColesterolYesenia Huaman RomeroAún no hay calificaciones
- ColesterolDocumento8 páginasColesterolJose Joel GonzalezAún no hay calificaciones
- ColesterolDocumento12 páginasColesterolJhon Fredy Carlos GomezAún no hay calificaciones
- ColesterolDocumento9 páginasColesterolDazamAún no hay calificaciones
- El Colesterol Es Un LípidoDocumento7 páginasEl Colesterol Es Un LípidoFerney CastroAún no hay calificaciones
- Seminario 12 - Texto - Metabolismo Del Colesterol - 2021Documento27 páginasSeminario 12 - Texto - Metabolismo Del Colesterol - 2021Joseph ReyesAún no hay calificaciones
- Bioquimica - ColesterolDocumento40 páginasBioquimica - ColesterolFernando Vazquez VarasAún no hay calificaciones
- Monografia de Colesterol y TrigliceridosDocumento23 páginasMonografia de Colesterol y TrigliceridosLiz M VillaVqz100% (1)
- Seminario 16 - Metabolismo Del Colesterol - 2020 PDFDocumento27 páginasSeminario 16 - Metabolismo Del Colesterol - 2020 PDFJorgeAún no hay calificaciones
- Colestero EquipoDocumento6 páginasColestero EquipoFrancisco Pérez José MarioAún no hay calificaciones
- Tema 16-Metabolismo Del ColesterolDocumento12 páginasTema 16-Metabolismo Del ColesterolLauraAún no hay calificaciones
- Metabolismo de ColesterolDocumento21 páginasMetabolismo de ColesterolJoel PMAún no hay calificaciones
- Trabajo FinalDocumento11 páginasTrabajo FinalFrancisth OrtizAún no hay calificaciones
- Clase 5. Sintesis ColesterolDocumento15 páginasClase 5. Sintesis ColesterolRONAL SANDOVAL TORRESAún no hay calificaciones
- Metabolismo Del ColesterolDocumento45 páginasMetabolismo Del ColesterolDiana VargasAún no hay calificaciones
- Sintesis de Acidos GrasosDocumento39 páginasSintesis de Acidos GrasosNancy Montserrat Villaseñor AranaAún no hay calificaciones
- Síntesis, Transporte y Excreción de ColesterolDocumento59 páginasSíntesis, Transporte y Excreción de ColesterolIsabel100% (1)
- Biosintesís de Colesterol y TriacilglicerolesDocumento25 páginasBiosintesís de Colesterol y TriacilglicerolesDeme Salazar KakoulidesAún no hay calificaciones
- ColesterolDocumento21 páginasColesterolArnoldo Jose Candanedo JaramilloAún no hay calificaciones
- Sintesis Del ColesterolDocumento22 páginasSintesis Del ColesterolIvanDeJesusPeñaGranielAún no hay calificaciones
- Metabolismo de Los IsoprenoidesDocumento15 páginasMetabolismo de Los IsoprenoidesAlejandra SarmientoAún no hay calificaciones
- Biosíntesis de Ácidos Grasos y Biosíntesis Del Colesterol.Documento22 páginasBiosíntesis de Ácidos Grasos y Biosíntesis Del Colesterol.Abrahamalberto HernandezAún no hay calificaciones
- 3.2 Metabolismo Del ColesterolDocumento31 páginas3.2 Metabolismo Del ColesterolNoelia LaraAún no hay calificaciones
- SESIÓN5. Síntesis de ColesterolDocumento23 páginasSESIÓN5. Síntesis de ColesterolDaniel MalpartidaAún no hay calificaciones
- Biosintesis Del Colesterol y Su Regulacion Descargar PDFDocumento1 páginaBiosintesis Del Colesterol y Su Regulacion Descargar PDFAaronAún no hay calificaciones
- Biosíntesis Del ColesterolDocumento19 páginasBiosíntesis Del ColesterolFrank RodríguezAún no hay calificaciones
- Bioquimica, Colesterol TotalDocumento3 páginasBioquimica, Colesterol TotalNatalia MaciasAún no hay calificaciones
- El ColesterolDocumento9 páginasEl ColesterolDavid TorresAún no hay calificaciones
- Bioquimica Metabolismo de La Prostaglandina y ColesterolDocumento9 páginasBioquimica Metabolismo de La Prostaglandina y Colesterolmarce bastidasAún no hay calificaciones
- Expo Biosintesis Del Colesterol OriginalDocumento10 páginasExpo Biosintesis Del Colesterol OriginalLuis AfgAún no hay calificaciones
- Metabolismo Del ColesterolDocumento52 páginasMetabolismo Del Colesterolfmh200950% (2)
- Metabolismos TAG y Los FosfolipidosDocumento55 páginasMetabolismos TAG y Los FosfolipidosDerian PoouAún no hay calificaciones
- Procesos Metabólicos DefDocumento33 páginasProcesos Metabólicos DefDiegoAlejandroCardonaAún no hay calificaciones
- Examen Final BIoquimica Teoria UnmsmDocumento8 páginasExamen Final BIoquimica Teoria UnmsmYASMIN MICHELLE MANRIQUE ALVAREZAún no hay calificaciones
- Biosintesis de Ácidos GrasosDocumento5 páginasBiosintesis de Ácidos GrasosbiologiaibericaAún no hay calificaciones
- TP Metabolismo de LipidosDocumento5 páginasTP Metabolismo de LipidosMili ReynaAún no hay calificaciones
- Biosíntesis Del ColesterolDocumento28 páginasBiosíntesis Del ColesterolEzequiel HernándezAún no hay calificaciones
- CarbohidratosDocumento2 páginasCarbohidratosclaudioAún no hay calificaciones
- Lipidos - Quimica I - 8. Biosintesis de ColesterolDocumento14 páginasLipidos - Quimica I - 8. Biosintesis de ColesterolYherson Ochoa LeónAún no hay calificaciones
- Biosíntesis Del ColesterolDocumento42 páginasBiosíntesis Del ColesterolLuz SaenzAún no hay calificaciones
- GlucolisisDocumento26 páginasGlucolisisJhanethMamaniAún no hay calificaciones
- Metabolismo Del Colesterol PDFDocumento9 páginasMetabolismo Del Colesterol PDFRuben Nuñez Quesada100% (2)
- Sintesis ColesterolDocumento20 páginasSintesis ColesterolAndrea PonceAún no hay calificaciones
- Metabolismo de Lipidos (Campbell & Farrell, Biochemistry, Capitulo 21)Documento37 páginasMetabolismo de Lipidos (Campbell & Farrell, Biochemistry, Capitulo 21)Eduardo DurazoAún no hay calificaciones
- Resumen Completo de La Biosíntesis de ColesterolDocumento2 páginasResumen Completo de La Biosíntesis de ColesterolCarolina DuranAún no hay calificaciones
- Final de Organica Esteroides..Documento34 páginasFinal de Organica Esteroides..Kleira Pérez Espinoza100% (1)
- Ciclo de Krebs, Cadena Respiratoria, Balance y Via de Las Pentosas. 2023Documento30 páginasCiclo de Krebs, Cadena Respiratoria, Balance y Via de Las Pentosas. 2023Lara VilteAún no hay calificaciones
- Trabajo Grupal - El ColesterolDocumento8 páginasTrabajo Grupal - El ColesterolYenniferAún no hay calificaciones
- Metabolismo Del ColesterolDocumento6 páginasMetabolismo Del ColesterolHILDA JHUVALY CHINOAPAZA BEJARAún no hay calificaciones
- Metabolismo de Lipidos IIDocumento10 páginasMetabolismo de Lipidos IIJose lorduyAún no hay calificaciones
- Metabolismo Del Colesterol y Cuerpos Cetonicos.Documento4 páginasMetabolismo Del Colesterol y Cuerpos Cetonicos.alguiensoy9512Aún no hay calificaciones
- Apa Quimica Sanguinea 4to Semestre 2020Documento23 páginasApa Quimica Sanguinea 4to Semestre 2020OSCAR CARDONA PANTOJAAún no hay calificaciones
- III Beta-Oxidacion y Cpos. CetonicosDocumento30 páginasIII Beta-Oxidacion y Cpos. CetonicosjijirijilloAún no hay calificaciones
- 6 Colesterol - Lipoproteinas 2Documento45 páginas6 Colesterol - Lipoproteinas 2ricardoAún no hay calificaciones
- Taller Lipidos 11960Documento8 páginasTaller Lipidos 11960Juan luis Guerra acostaAún no hay calificaciones
- Marco TeoricoDocumento8 páginasMarco TeoricoAbby RamirezAún no hay calificaciones
- CarbohidratosDocumento18 páginasCarbohidratosSilva Arellano AxelAún no hay calificaciones
- Estructura y Composición de Los Aceites y Grasas PDFDocumento106 páginasEstructura y Composición de Los Aceites y Grasas PDFDAHANA CAROLINA VARGAS AGROTAAún no hay calificaciones
- Autoevaluación Lípidos IDocumento3 páginasAutoevaluación Lípidos ICeciliaAún no hay calificaciones
- MONOGRAFIA Perfil HepaticoDocumento21 páginasMONOGRAFIA Perfil HepaticoWilly Fabio Flores0% (1)
- 4.10 Función de Transporte de Lípidos en La Sangre, Lipoproteínas, Estructura y Función de Quilomicrones, VLDL, LDL y HDL.Documento14 páginas4.10 Función de Transporte de Lípidos en La Sangre, Lipoproteínas, Estructura y Función de Quilomicrones, VLDL, LDL y HDL.Ilian Guzmán MendozaAún no hay calificaciones
- Practica 9 TrgliceridosDocumento14 páginasPractica 9 TrgliceridosFiorella Rios GoñyAún no hay calificaciones
- Caso 2Documento5 páginasCaso 2Diego HuertasAún no hay calificaciones
- Bioquimica 2-TEMA 12Documento21 páginasBioquimica 2-TEMA 12ArlettA.MendozaAún no hay calificaciones
- CLASE 1 Lipidos. - DigestionDocumento36 páginasCLASE 1 Lipidos. - DigestionCarlos HfAún no hay calificaciones
- Determinacion de Lipidos en El Laboratorio.Documento7 páginasDeterminacion de Lipidos en El Laboratorio.Montserrat HerreraAún no hay calificaciones
- Formacion de Tapon PlaquetarioDocumento21 páginasFormacion de Tapon PlaquetarioYave Aro Rodriguez100% (1)
- Los Lipidos-Cuestionario Unidad 4Documento12 páginasLos Lipidos-Cuestionario Unidad 4Michelle GrandeAún no hay calificaciones
- LipidogramaDocumento6 páginasLipidogramaNajari RijanaAún no hay calificaciones
- Lípidos Tema Nº3Documento31 páginasLípidos Tema Nº3Gowther DelgadilloAún no hay calificaciones
- 2 Maynar-Presentacion DislipidemiaDocumento17 páginas2 Maynar-Presentacion Dislipidemiajulialunagonzalez98Aún no hay calificaciones
- El Aguacate-2212029208Documento5 páginasEl Aguacate-2212029208Luisa HuertasAún no hay calificaciones
- Gmbii 2023 V1 12 01 23Documento105 páginasGmbii 2023 V1 12 01 23Maricarmen OsorioAún no hay calificaciones
- Clase Semana 3 PDFDocumento41 páginasClase Semana 3 PDFLIZ ANGIE URBANO ESPIRITUAún no hay calificaciones
- BQ 17 Chi 7 Metabolismo de Lipidos HeliDocumento55 páginasBQ 17 Chi 7 Metabolismo de Lipidos HeliDaniel Vasquez SandovalAún no hay calificaciones
- Castellano CuentoDocumento23 páginasCastellano CuentoDaniela JiménezAún no hay calificaciones
- Metabolismo de Los LípidosDocumento26 páginasMetabolismo de Los LípidosDiara MaylénAún no hay calificaciones
- LipidosDocumento11 páginasLipidosGabriela VegaAún no hay calificaciones
- Solano Aguilar Diego ArmandoDocumento7 páginasSolano Aguilar Diego ArmandoNicole Dayana Patiño SerranoAún no hay calificaciones
- LípidosDocumento18 páginasLípidosMario Ernesto AlvarengaAún no hay calificaciones
- LaboratorioDocumento1 páginaLaboratorioJose EliasAún no hay calificaciones
- Informe LipoproteinasDocumento4 páginasInforme LipoproteinasJorge Andres EspinozaAún no hay calificaciones
- Guia LipoproteinasDocumento8 páginasGuia LipoproteinasManuel de Jesus100% (1)
- Dislipidemia y Sindrome MetabolicoDocumento49 páginasDislipidemia y Sindrome MetabolicoCamilo Jose Lopez MolinaAún no hay calificaciones