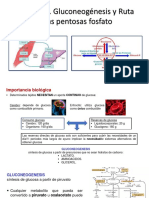
También podría gustarte
- Tema 17Documento32 páginasTema 17CERBEROSHIVAAún no hay calificaciones
- Ayudas ergogénicas y nutricionalesDe EverandAyudas ergogénicas y nutricionalesCalificación: 5 de 5 estrellas5/5 (2)
- Gluconeogenesis 1Documento34 páginasGluconeogenesis 1Ernesto GarciaAún no hay calificaciones
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- Gluconeogenesis 2 Doparcial Bioqui 2023 PDFDocumento28 páginasGluconeogenesis 2 Doparcial Bioqui 2023 PDFDebora RuizAún no hay calificaciones
- Ciclo de KrebsDocumento85 páginasCiclo de KrebsDANNA VANESSA FLOREZ RUEDAAún no hay calificaciones
- Sesion 05 Glicólisis 2s2023Documento34 páginasSesion 05 Glicólisis 2s2023ola467442Aún no hay calificaciones
- Metabolismo de CarbohidratosDocumento33 páginasMetabolismo de CarbohidratosXiomara Sanchez CahuanaAún no hay calificaciones
- GluconeogénesisDocumento3 páginasGluconeogénesisM PAún no hay calificaciones
- Metabolismo de CarbohidratosDocumento73 páginasMetabolismo de CarbohidratosGrecia Vera VallejosAún no hay calificaciones
- Bioquímica Gluneogenesis y GlugenolisisDocumento104 páginasBioquímica Gluneogenesis y GlugenolisisDANNA VANESSA FLOREZ RUEDAAún no hay calificaciones
- GLUCONEOGENESISDocumento32 páginasGLUCONEOGENESISzatanna66100% (2)
- 3.1 GluconeogensisDocumento18 páginas3.1 GluconeogensisAlfredo PalaciosAún no hay calificaciones
- UNIDAD IV B - Gluconeoge - Nesis - 21 Abril2016Documento30 páginasUNIDAD IV B - Gluconeoge - Nesis - 21 Abril2016Natalia Paz Solar MartinezAún no hay calificaciones
- Gluconeogénesis, Glucogenóli Sis, Glucogénesis, Via de Las Pentosa FosfatoDocumento43 páginasGluconeogénesis, Glucogenóli Sis, Glucogénesis, Via de Las Pentosa FosfatoBertha GuivinAún no hay calificaciones
- 7) Los Carbohidratos en La Nutrición (2da Parte) Pregrad.Documento43 páginas7) Los Carbohidratos en La Nutrición (2da Parte) Pregrad.Cindy Ely Lucero Quispe AndradeAún no hay calificaciones
- Rutas Metabolicas CHOSDocumento51 páginasRutas Metabolicas CHOSAlfredo GonzalezAún no hay calificaciones
- Semana 3Documento70 páginasSemana 3Thamara Cruz100% (1)
- Seminario Gluconeogénesis RealizadoDocumento6 páginasSeminario Gluconeogénesis RealizadoLuz Jasmine FigueroaAún no hay calificaciones
- CarbohidratosDocumento30 páginasCarbohidratosTrash talkingAún no hay calificaciones
- Respiración celular: la conversión de energía en ATPDocumento16 páginasRespiración celular: la conversión de energía en ATPYanet Lucia REYES FEBREAún no hay calificaciones
- 12 - Glucolisis y Gluconeogenesis-2022Documento47 páginas12 - Glucolisis y Gluconeogenesis-2022Maximiliano MonsalveAún no hay calificaciones
- Gluconeogénesisglucogenólisisglucogénesisvia de Las Pentosa Fosfato 3Documento43 páginasGluconeogénesisglucogenólisisglucogénesisvia de Las Pentosa Fosfato 3Pilar Polo GonzalezAún no hay calificaciones
- Gluconeogénesis - Ciclo de Las PentosasDocumento57 páginasGluconeogénesis - Ciclo de Las PentosasUSMP FN ARCHIVOSAún no hay calificaciones
- GluconeogenesisDocumento37 páginasGluconeogenesisStefanny Gabriela100% (1)
- SEMANA 8. Gluconeogénesis y Ruta de Las Pentosas FosfatoDocumento44 páginasSEMANA 8. Gluconeogénesis y Ruta de Las Pentosas FosfatoVIVIANA ZULEMA AZA�ERO MAGANAún no hay calificaciones
- Catabolismo glucosaDocumento3 páginasCatabolismo glucosanataliAún no hay calificaciones
- Expo Bioquimica Metabolismo de Carbohidratos 2023Documento24 páginasExpo Bioquimica Metabolismo de Carbohidratos 2023Jhonny Alejandro Chadid BenítezAún no hay calificaciones
- Factores Físico Químicos en La Producción Del BiogásDocumento63 páginasFactores Físico Químicos en La Producción Del BiogásPANDITA KAWAI M.Aún no hay calificaciones
- 9 Metabolismo Celular2Documento78 páginas9 Metabolismo Celular2Agustín MarceroAún no hay calificaciones
- 09 Glicolisis-FermentacionDocumento49 páginas09 Glicolisis-FermentacionRomina SalazarAún no hay calificaciones
- GluconeogenesisDocumento24 páginasGluconeogenesisEloy Alfonso Flores CastilloAún no hay calificaciones
- RUTAS DE LA GLUCOSA A NIVEL CELULAR Pablo HernándezDocumento9 páginasRUTAS DE LA GLUCOSA A NIVEL CELULAR Pablo HernándezPablo HernandezAún no hay calificaciones
- Esquema de Catabolismo de Los CarbohidratosDocumento5 páginasEsquema de Catabolismo de Los CarbohidratosJuanPérezAún no hay calificaciones
- Resumen Tema 6.2Documento15 páginasResumen Tema 6.2juanmideluxeAún no hay calificaciones
- Tema 2. CarbohidratosDocumento22 páginasTema 2. CarbohidratosPaola Ojeda FernándezAún no hay calificaciones
- Clase 2-Metabolismo de CarbohidratosDocumento41 páginasClase 2-Metabolismo de CarbohidratosOrions MTovalinoAún no hay calificaciones
- 1 - Introducción Al Metabolismo de La Glucosa-GlucolisisDocumento71 páginas1 - Introducción Al Metabolismo de La Glucosa-GlucolisisGabi RochaAún no hay calificaciones
- GluconeogénesisDocumento58 páginasGluconeogénesisFeer RangelAún no hay calificaciones
- Metabolismo de Carbohidratos I Per 2022Documento35 páginasMetabolismo de Carbohidratos I Per 2022bryan lovoAún no hay calificaciones
- 1.3 CarbohidratosDocumento39 páginas1.3 Carbohidratospatrick floresAún no hay calificaciones
- Metabolismo de Los CarbohidratosDocumento37 páginasMetabolismo de Los CarbohidratosKARLA GONZALEZ QUIROZ100% (1)
- Apuntes de GlucólisisDocumento8 páginasApuntes de GlucólisisIsba Shadai Estrada GarciaAún no hay calificaciones
- Gluconeogénesis y control glucosaDocumento6 páginasGluconeogénesis y control glucosaIsba Shadai Estrada GarciaAún no hay calificaciones
- MetabolismoCHO (B)Documento48 páginasMetabolismoCHO (B)SarahAún no hay calificaciones
- Catabolismo GlucidosDocumento3 páginasCatabolismo GlucidosAna Garcia MartínezAún no hay calificaciones
- Macronutrientes SSDocumento69 páginasMacronutrientes SSJULIETAAún no hay calificaciones
- MN 2Documento3 páginasMN 2CristAmuert 3Aún no hay calificaciones
- Metabolismo carbohidratos glucosa glucólisis ciclo krebsDocumento97 páginasMetabolismo carbohidratos glucosa glucólisis ciclo krebsJacqueline Hernandez HernandezAún no hay calificaciones
- Metabolismo de CarbohidratosDocumento52 páginasMetabolismo de CarbohidratosLauraSofiaRamirezAún no hay calificaciones
- Metabolismo de Hidratos de Carbono Parte 1Documento61 páginasMetabolismo de Hidratos de Carbono Parte 1Mundo VacioAún no hay calificaciones
- Clase Glucolisis.Documento42 páginasClase Glucolisis.les12michuAún no hay calificaciones
- Bioquimica RepasoDocumento13 páginasBioquimica RepasoholaAún no hay calificaciones
- Gluconeogénesis: Síntesis de glucosa a partir de otros sustratosDocumento18 páginasGluconeogénesis: Síntesis de glucosa a partir de otros sustratosCarlota GuardiaAún no hay calificaciones
- Metabolismo de Los CarbohidratosDocumento53 páginasMetabolismo de Los CarbohidratosErick Andrés Herbas García50% (2)
- Metabolismo de Los CarbohidratosDocumento31 páginasMetabolismo de Los CarbohidratosIdaniaAún no hay calificaciones
- Metabolismo de CarbohidratosDocumento44 páginasMetabolismo de CarbohidratosJesiel PadovaniAún no hay calificaciones
- Gluconeogenesis PDFDocumento19 páginasGluconeogenesis PDFMeli Saavedra100% (1)
- Tarea1 Unidad4Documento12 páginasTarea1 Unidad4Abrahamalberto HernandezAún no hay calificaciones
- CLASE 10 Gases Disueltos CO2Documento8 páginasCLASE 10 Gases Disueltos CO2carloscaunaAún no hay calificaciones
- Gases disueltos en lagos y factores que afectan su solubilidadDocumento13 páginasGases disueltos en lagos y factores que afectan su solubilidadKIMBERLY MARY ARIAS PINEDAAún no hay calificaciones
- Tema 2 10 Dias de OracionDocumento8 páginasTema 2 10 Dias de OracionKIMBERLY MARY ARIAS PINEDAAún no hay calificaciones
- CLASE 10 Aspectos Químicos de Las Aguas (Limnologia Quimica)Documento4 páginasCLASE 10 Aspectos Químicos de Las Aguas (Limnologia Quimica)KIMBERLY MARY ARIAS PINEDAAún no hay calificaciones
- El Ciclo de La Urea Comienza en El Interior de Las Mitocondrias de Los HepatocitosDocumento1 páginaEl Ciclo de La Urea Comienza en El Interior de Las Mitocondrias de Los HepatocitosKIMBERLY MARY ARIAS PINEDAAún no hay calificaciones
- Tema 4 10 Dias de OracionDocumento8 páginasTema 4 10 Dias de OracionKIMBERLY MARY ARIAS PINEDAAún no hay calificaciones
- Tema 4 10 Dias de OracionDocumento8 páginasTema 4 10 Dias de OracionKIMBERLY MARY ARIAS PINEDAAún no hay calificaciones
- Ejercicio Teoria 2Documento1 páginaEjercicio Teoria 2KIMBERLY MARY ARIAS PINEDAAún no hay calificaciones
- PoliticaDocumento1 páginaPoliticaKIMBERLY MARY ARIAS PINEDAAún no hay calificaciones
- El futuro revelado: descubriendo la verdadDocumento11 páginasEl futuro revelado: descubriendo la verdadKIMBERLY MARY ARIAS PINEDAAún no hay calificaciones
- Escala de GrisesDocumento1 páginaEscala de GrisesKIMBERLY MARY ARIAS PINEDAAún no hay calificaciones
- La Educación en El TahuantinsuyoDocumento2 páginasLa Educación en El TahuantinsuyoKIMBERLY MARY ARIAS PINEDAAún no hay calificaciones
- Practica Nº12. Auditoria Haccp AcuiculturaDocumento38 páginasPractica Nº12. Auditoria Haccp AcuiculturaKIMBERLY MARY ARIAS PINEDAAún no hay calificaciones
- Informe - Aceite - DestilacionDocumento14 páginasInforme - Aceite - Destilaciondany condori crispinAún no hay calificaciones
- ProblemaDocumento2 páginasProblemaEmmanuel Escobar AcostaAún no hay calificaciones
- MC-Caja de Valvula Desde CONSIDERACIONES DE DISEÑO 3Documento18 páginasMC-Caja de Valvula Desde CONSIDERACIONES DE DISEÑO 3gerardoAún no hay calificaciones
- Diseño de mezclas de hormigónDocumento45 páginasDiseño de mezclas de hormigónIran GreciaAún no hay calificaciones
- Proceso Productivo para La Obtención Del Bioetanol A Partir Del Bagazo de Caña de AzúcarDocumento6 páginasProceso Productivo para La Obtención Del Bioetanol A Partir Del Bagazo de Caña de AzúcarxShugoSx delongeAún no hay calificaciones
- Laboratorio Operaciones QuímicasDocumento4 páginasLaboratorio Operaciones QuímicasMARCK ANTHONY ALVAREZ POLANCOAún no hay calificaciones
- Edusol 1789-8091: E-Issn: Edusol@Cug - Co.CuDocumento12 páginasEdusol 1789-8091: E-Issn: Edusol@Cug - Co.CubernyrbAún no hay calificaciones
- Tratado de Criminalistica Tomo I PDFDocumento370 páginasTratado de Criminalistica Tomo I PDFcarlaAún no hay calificaciones
- Cromatografía Pigmentos VegetalesDocumento23 páginasCromatografía Pigmentos Vegetalesmaysilee-katnissAún no hay calificaciones
- Instituto Politécnico Nacional: Escuela Superior de Ingeniería Química E Industrias ExtractivasDocumento117 páginasInstituto Politécnico Nacional: Escuela Superior de Ingeniería Química E Industrias ExtractivaspabloAún no hay calificaciones
- PRÁCTICA 3. Determinación Espectrofotométrica de Hierro en Formas Farmacéuticas.Documento8 páginasPRÁCTICA 3. Determinación Espectrofotométrica de Hierro en Formas Farmacéuticas.Edith A CamposAún no hay calificaciones
- Ficha Termometro DigitalDocumento1 páginaFicha Termometro DigitalsermedAún no hay calificaciones
- Laboratorio 1-2Documento26 páginasLaboratorio 1-2luis_pqm1Aún no hay calificaciones
- II.3.TECNICAS DE SIEMBRA (Autoguardado)Documento15 páginasII.3.TECNICAS DE SIEMBRA (Autoguardado)Elvis Giovanni Cruz VenturaAún no hay calificaciones
- Fuerza y Calculo de ph-pOHDocumento16 páginasFuerza y Calculo de ph-pOHIlse HernándezAún no hay calificaciones
- Propiedades y Rombos de SeguridadDocumento2 páginasPropiedades y Rombos de SeguridadLa Ruta del CaminanteAún no hay calificaciones
- 1S. HibridaciónDocumento15 páginas1S. HibridaciónDeysi ChavezAún no hay calificaciones
- Aldehidos y CetonasDocumento4 páginasAldehidos y CetonasMariel CossioAún no hay calificaciones
- TRABAJO PROC. WPS CorregidoDocumento42 páginasTRABAJO PROC. WPS Corregidoeduardo alata ormeñoAún no hay calificaciones
- Practica 4 CorrosiónDocumento14 páginasPractica 4 CorrosiónVictor RamirezAún no hay calificaciones
- P5_Ensayos_Reactivos_Nitrato_CericoDocumento3 páginasP5_Ensayos_Reactivos_Nitrato_CericoDANIELA ESCOBAR MAHECHAAún no hay calificaciones
- Semana 10 HISTORIA. DIAZ - Jose Luis Diaz TurpoDocumento2 páginasSemana 10 HISTORIA. DIAZ - Jose Luis Diaz Turpoyedy garid sotoAún no hay calificaciones
- Historia de La BioquímicaDocumento2 páginasHistoria de La Bioquímicajohanna cajilimaAún no hay calificaciones
- ZN03-PV. EPOXY ZINC PRIMER QD (Abril 2016)Documento2 páginasZN03-PV. EPOXY ZINC PRIMER QD (Abril 2016)ANIBALLOPEZVEGAAún no hay calificaciones
- Practica 1 Microbiologia Predictiva DesaDocumento11 páginasPractica 1 Microbiologia Predictiva Desakaren lhzAún no hay calificaciones
- Curso para Operadores de CalderasDocumento74 páginasCurso para Operadores de Calderasmantencion CRIAún no hay calificaciones
- Modulo2 Practica2Documento10 páginasModulo2 Practica2Esteban CadenaAún no hay calificaciones
- Examen Parcial #01 de Hidrologia 30.09.2022Documento9 páginasExamen Parcial #01 de Hidrologia 30.09.2022Jean CarlosAún no hay calificaciones
- MANUAL PILOTES METALICOS Lr-MinDocumento68 páginasMANUAL PILOTES METALICOS Lr-MiningetabascoAún no hay calificaciones
- Inyección de PlásticoDocumento18 páginasInyección de PlásticoMario HinostrozaAún no hay calificaciones
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- La Dieta Antiinflamatoria: Haz estos cambios simples y económicos en tu dieta y comienza a sentirte mejor dentro de 24 horas! (Spanish Edition)De EverandLa Dieta Antiinflamatoria: Haz estos cambios simples y económicos en tu dieta y comienza a sentirte mejor dentro de 24 horas! (Spanish Edition)Calificación: 5 de 5 estrellas5/5 (4)
- Detox: Alimentación depurativa para tu salud: Dietas, zumos, batidos y recetas para depurar tu cuerpo de forma naturalDe EverandDetox: Alimentación depurativa para tu salud: Dietas, zumos, batidos y recetas para depurar tu cuerpo de forma naturalCalificación: 5 de 5 estrellas5/5 (7)
- 50 Recetas Ketodeliciosas, Libro De Recetas Para Dieta CetogénicaDe Everand50 Recetas Ketodeliciosas, Libro De Recetas Para Dieta CetogénicaCalificación: 5 de 5 estrellas5/5 (3)
- Recetas para Diabéticos y Alimentos que Controlan la Diabetes: Plan de Comidas para Diabéticos de todas las edades que deseen una Dieta Saludable: Plan de comidas para diabéticos de todas las edades que deseen una dieta saludableDe EverandRecetas para Diabéticos y Alimentos que Controlan la Diabetes: Plan de Comidas para Diabéticos de todas las edades que deseen una Dieta Saludable: Plan de comidas para diabéticos de todas las edades que deseen una dieta saludableCalificación: 5 de 5 estrellas5/5 (1)
- Dieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasDe EverandDieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasCalificación: 4 de 5 estrellas4/5 (9)
- Plan De Dieta Para La Resistencia A La Insulina & Sistema Inmunológico En EspañolDe EverandPlan De Dieta Para La Resistencia A La Insulina & Sistema Inmunológico En EspañolCalificación: 3.5 de 5 estrellas3.5/5 (2)
- Mega Piernas: Entrenamiento Científico de PiernasDe EverandMega Piernas: Entrenamiento Científico de PiernasCalificación: 4 de 5 estrellas4/5 (4)
- CLEAN 7 \ El Método Clean 7 (Spanish edition): Detoxifica y rejuvenece tu cuerpo en tan sólo siete díasDe EverandCLEAN 7 \ El Método Clean 7 (Spanish edition): Detoxifica y rejuvenece tu cuerpo en tan sólo siete díasCalificación: 5 de 5 estrellas5/5 (1)
- Cómo Desintoxicar el Cuerpo de Metales Pesados: El Secreto Poco Hablado para Sentirte Mejor, Bajar de Peso y Tener más EnergíaDe EverandCómo Desintoxicar el Cuerpo de Metales Pesados: El Secreto Poco Hablado para Sentirte Mejor, Bajar de Peso y Tener más EnergíaCalificación: 4.5 de 5 estrellas4.5/5 (2)
- ¡Sana tu intestino!: Recetario de la Dieta GAPSDe Everand¡Sana tu intestino!: Recetario de la Dieta GAPSCalificación: 4.5 de 5 estrellas4.5/5 (4)
- Aprender a comer solo: Manual sobre el método Baby Led WeaningDe EverandAprender a comer solo: Manual sobre el método Baby Led WeaningCalificación: 4.5 de 5 estrellas4.5/5 (2)
- Postres Saludables para Disfrutar sin Culpa. Postres Paleo sin Gluten, Azucar ni LactosaDe EverandPostres Saludables para Disfrutar sin Culpa. Postres Paleo sin Gluten, Azucar ni LactosaCalificación: 5 de 5 estrellas5/5 (17)
- Sana tu Cuerpo, Calma tu Mente: Desintoxicar Hígado, Intestino Permeable, Salud Hormonal, Curación Emocional, Relajación, Ansiedad y Sanidad mental, Atención Plena, Psicoterapia y NutriciónDe EverandSana tu Cuerpo, Calma tu Mente: Desintoxicar Hígado, Intestino Permeable, Salud Hormonal, Curación Emocional, Relajación, Ansiedad y Sanidad mental, Atención Plena, Psicoterapia y NutriciónCalificación: 5 de 5 estrellas5/5 (4)
- Mente sana, cuerpo sano: El poder de una mentalidad saludableDe EverandMente sana, cuerpo sano: El poder de una mentalidad saludableCalificación: 5 de 5 estrellas5/5 (90)
- El libro de la nutrición práctica: Un vademécum completo y riguroso sobre la alimentación y cómo influye en su saludDe EverandEl libro de la nutrición práctica: Un vademécum completo y riguroso sobre la alimentación y cómo influye en su saludCalificación: 4 de 5 estrellas4/5 (1)
- Mega Rutinas De Entrenamiento Con Peso CorporalDe EverandMega Rutinas De Entrenamiento Con Peso CorporalCalificación: 3 de 5 estrellas3/5 (2)
- Manual de nutrición deportiva (Color)De EverandManual de nutrición deportiva (Color)Calificación: 4 de 5 estrellas4/5 (21)
- Ayuno Intermitente: ¡Pierde Peso, Sana Tu Cuerpo y Vive una Vida Saludable!De EverandAyuno Intermitente: ¡Pierde Peso, Sana Tu Cuerpo y Vive una Vida Saludable!Calificación: 3.5 de 5 estrellas3.5/5 (25)
- Cómo curar la artritis: Curación natural de la artritis, la artrosis, la gota y la fibromialgiaDe EverandCómo curar la artritis: Curación natural de la artritis, la artrosis, la gota y la fibromialgiaCalificación: 5 de 5 estrellas5/5 (17)
- El ayuno como estilo de vida: Descubre las claves para incorporar el ayuno intermitente a tu vida y disfrutar de sus beneficios para la salud y la pérdida de pesoDe EverandEl ayuno como estilo de vida: Descubre las claves para incorporar el ayuno intermitente a tu vida y disfrutar de sus beneficios para la salud y la pérdida de pesoCalificación: 5 de 5 estrellas5/5 (10)