También podría gustarte
- CARBOHIDRATOSDocumento90 páginasCARBOHIDRATOSAnnel ValenzuelaAún no hay calificaciones
- Metabolismo de Hemoglobina (DV)Documento9 páginasMetabolismo de Hemoglobina (DV)Dary Deivy Varela AracheAún no hay calificaciones
- TequilaDocumento4 páginasTequilaAnonymous LnLs17uVAún no hay calificaciones
- Resina Urea FormaldehidoDocumento13 páginasResina Urea FormaldehidoCiro Cayllahua EscalanteAún no hay calificaciones
- EJERCICIOS - AMINOÁCIDOS, PÉPTIDOS y PROTEÍNASDocumento2 páginasEJERCICIOS - AMINOÁCIDOS, PÉPTIDOS y PROTEÍNASZuilen Robles Rodriguez100% (1)
- Metodo BiuretDocumento9 páginasMetodo BiuretPablo CuevasAún no hay calificaciones
- Lactato DeshidrogenasaDocumento2 páginasLactato DeshidrogenasaCarlos SegovianoAún no hay calificaciones
- Espectrofotometria Acido AscorbicoDocumento5 páginasEspectrofotometria Acido AscorbicorogerAún no hay calificaciones
- Reporte Proteinas, CONCENTRACION DE PROTEINASDocumento3 páginasReporte Proteinas, CONCENTRACION DE PROTEINASletyss100% (1)
- TriglicéridosDocumento7 páginasTriglicéridosSelmerAún no hay calificaciones
- Unidad 7 Integracin MetabolicaDocumento114 páginasUnidad 7 Integracin MetabolicaPedro Miranda Aponte86% (7)
- AlbúminaDocumento3 páginasAlbúminaMelanie SamurioAún no hay calificaciones
- Metabolismo de Los CarbohidratosDocumento30 páginasMetabolismo de Los CarbohidratosSandy GonzalezAún no hay calificaciones
- Metabolismo de Los GlúcidosDocumento18 páginasMetabolismo de Los GlúcidosstevemburgAún no hay calificaciones
- Metabolismo de Acidos Grasos.Documento7 páginasMetabolismo de Acidos Grasos.Mariel AybarAún no hay calificaciones
- Semana 9. Metabolismo de Los Ácidos Grasos, Biosíntesis de Fosfolípidos, Glicólipidos, Colesterol.Documento69 páginasSemana 9. Metabolismo de Los Ácidos Grasos, Biosíntesis de Fosfolípidos, Glicólipidos, Colesterol.Carmen Villegas GermànAún no hay calificaciones
- GLUCONEOGÉNESISDocumento14 páginasGLUCONEOGÉNESISnamames jodeteAún no hay calificaciones
- Metabolismo y Excreción de Farmacos (Informe)Documento9 páginasMetabolismo y Excreción de Farmacos (Informe)FalconHC100% (2)
- 24 Metabolismo LípidosDocumento31 páginas24 Metabolismo LípidosClau VargasAún no hay calificaciones
- Bioquímica Metabolismo de ProteinasDocumento11 páginasBioquímica Metabolismo de ProteinasRafael ColmenaresAún no hay calificaciones
- Metabolismo de Los Carbohidratos 1Documento5 páginasMetabolismo de Los Carbohidratos 1Aran VeraAún no hay calificaciones
- Determinación de Colesterol TotalDocumento3 páginasDeterminación de Colesterol TotalAna Teresa Luna OliveraAún no hay calificaciones
- CARAMELIZACIONDocumento1 páginaCARAMELIZACIONMarco JuniorAún no hay calificaciones
- Fibra Dietetica InformeDocumento11 páginasFibra Dietetica InformeJohnAndresPinargoteAún no hay calificaciones
- MalonatoDocumento4 páginasMalonatoDavid GuachaminAún no hay calificaciones
- Ciclo de La UreaDocumento65 páginasCiclo de La UreaAlexandraMedinaCastilloAún no hay calificaciones
- Secresion SalivalDocumento46 páginasSecresion Salivalcoolboy17Aún no hay calificaciones
- Tipos de Agua XDDocumento1 páginaTipos de Agua XDANDRES FELIPE DIAZ SANCHEZAún no hay calificaciones
- Calidad de Los AlimentosDocumento71 páginasCalidad de Los AlimentosJohnny SalazarAún no hay calificaciones
- Práctica 3 Digestión de La Albúmina Por Pepsina Industrial.Documento7 páginasPráctica 3 Digestión de La Albúmina Por Pepsina Industrial.Glendiux_vaneAún no hay calificaciones
- Determinación de Humedad 1Documento4 páginasDeterminación de Humedad 1Cesia Fuentes100% (1)
- Los Carbohidratos Ensayo MioDocumento4 páginasLos Carbohidratos Ensayo MioNicole AndradeAún no hay calificaciones
- Formacion y Transporte de AmoniacoDocumento4 páginasFormacion y Transporte de AmoniacoMariela MaldonadoAún no hay calificaciones
- ProteinasDocumento14 páginasProteinasFabián Vasco GuerreroAún no hay calificaciones
- Salmonella Typhi en AlimentosDocumento59 páginasSalmonella Typhi en Alimentosshirley kinderly vargas archemtiAún no hay calificaciones
- Analisis de Leche CrudaDocumento9 páginasAnalisis de Leche CrudaJhonathan Paul Becerra VilelaAún no hay calificaciones
- PRACTICA 2 Curva de Tolerancia A La GlucosaDocumento6 páginasPRACTICA 2 Curva de Tolerancia A La GlucosaalexisAún no hay calificaciones
- Acidos Grasos 2Documento27 páginasAcidos Grasos 2Aleli Chinguel CastilloAún no hay calificaciones
- Metodos Lactosa PDFDocumento146 páginasMetodos Lactosa PDFChristopher Gamble100% (2)
- Metabolismo Catabolismo y AnabolismoDocumento16 páginasMetabolismo Catabolismo y AnabolismoCARLOS MANRIQUEAún no hay calificaciones
- Laboratorio de Aminoácidos y ProteínasDocumento8 páginasLaboratorio de Aminoácidos y ProteínasKaren RestrepoAún no hay calificaciones
- 6 - Metabolismo LipídicoDocumento27 páginas6 - Metabolismo LipídicoJosh ReyAún no hay calificaciones
- INFORME10Documento10 páginasINFORME10Nicole GuzmanAún no hay calificaciones
- Practica 6 Analisis de Vitaminas y MineralesDocumento5 páginasPractica 6 Analisis de Vitaminas y MineralesLuis BahachilleAún no hay calificaciones
- Seminario. Glucolisis y Oxidacion Del Piruvato.Documento45 páginasSeminario. Glucolisis y Oxidacion Del Piruvato.lorena navarreteAún no hay calificaciones
- Efecto de La Adrenalina y La InsulinaDocumento9 páginasEfecto de La Adrenalina y La InsulinaJavier YactayoAún no hay calificaciones
- Catabolismo de AminoacidosDocumento20 páginasCatabolismo de AminoacidosXimena Véliz DaudAún no hay calificaciones
- Trabajo de Bioquimica Del YogurtDocumento22 páginasTrabajo de Bioquimica Del Yogurtvalerio0% (1)
- HEMOPROTEINASDocumento2 páginasHEMOPROTEINASVale DegraveAún no hay calificaciones
- Conservación y Almacenamiento de AlimentosDocumento5 páginasConservación y Almacenamiento de AlimentoslauralorenaleongomezAún no hay calificaciones
- Glu PapaDocumento5 páginasGlu PapaDavid100% (1)
- Catabolismo de Los AminoácidosDocumento39 páginasCatabolismo de Los AminoácidosEd BozaAún no hay calificaciones
- 2 Clasificación de Los Aminoácidos en Función de La Naturaleza de Su Cadena LateralDocumento4 páginas2 Clasificación de Los Aminoácidos en Función de La Naturaleza de Su Cadena LateralAsier SantamaríaAún no hay calificaciones
- Determinación Calórico de Los AlimentosDocumento18 páginasDeterminación Calórico de Los AlimentosFrankling Jarly Shapiama Tafur100% (1)
- Carotenoides 2011Documento31 páginasCarotenoides 2011Ruben Kapa TiconaAún no hay calificaciones
- Practica Bromatologia 2019 (Ultima)Documento59 páginasPractica Bromatologia 2019 (Ultima)mauricioAún no hay calificaciones
- 11 Actividad EnzimaticaDocumento6 páginas11 Actividad EnzimaticaRaquel Yoselin Guzman RosasAún no hay calificaciones
- Observacion Directa MicoorganismosDocumento2 páginasObservacion Directa MicoorganismosGabriela Pasion BazanAún no hay calificaciones
- Alteraciones de GlucosaDocumento7 páginasAlteraciones de GlucosaAlexRichardRichardItamariAún no hay calificaciones
- Enzimas PeptidasasDocumento26 páginasEnzimas PeptidasasJavier Lambraño CastilloAún no hay calificaciones
- GluconeogenesisDocumento20 páginasGluconeogenesisEsperanza De JesusAún no hay calificaciones
- GlucoDocumento20 páginasGlucoBetty Javier GradosAún no hay calificaciones
- Gluconeogenesis 1Documento34 páginasGluconeogenesis 1Ernesto GarciaAún no hay calificaciones
- Retie Pre-InformeDocumento3 páginasRetie Pre-InformeDANNA VANESSA FLOREZ RUEDAAún no hay calificaciones
- Analisis CotizacionesDocumento3 páginasAnalisis CotizacionesDANNA VANESSA FLOREZ RUEDAAún no hay calificaciones
- Imagenes de La Guía de Manejo RápidoDocumento2 páginasImagenes de La Guía de Manejo RápidoDANNA VANESSA FLOREZ RUEDAAún no hay calificaciones
- FORMATO HOJA DE VIDA pHmETRODocumento22 páginasFORMATO HOJA DE VIDA pHmETRODANNA VANESSA FLOREZ RUEDAAún no hay calificaciones
- Lab 2 - PRESENCIAL Diodo Zener y LedDocumento8 páginasLab 2 - PRESENCIAL Diodo Zener y LedDANNA VANESSA FLOREZ RUEDAAún no hay calificaciones
- LAB 1 - METODOLOGÍA EN V - Martes4pmDocumento2 páginasLAB 1 - METODOLOGÍA EN V - Martes4pmDANNA VANESSA FLOREZ RUEDAAún no hay calificaciones
- Taller de VITAMINAS Y COENZIMASDocumento5 páginasTaller de VITAMINAS Y COENZIMASDANNA VANESSA FLOREZ RUEDAAún no hay calificaciones
- Ciclo de CarnotDocumento4 páginasCiclo de CarnotDANNA VANESSA FLOREZ RUEDAAún no hay calificaciones
- Base de Datos SonarDocumento17 páginasBase de Datos SonarDANNA VANESSA FLOREZ RUEDA100% (1)
- Ciclo de KrebsDocumento85 páginasCiclo de KrebsDANNA VANESSA FLOREZ RUEDAAún no hay calificaciones
- ESTROGENODocumento6 páginasESTROGENODANNA VANESSA FLOREZ RUEDAAún no hay calificaciones
- Goma FinalDocumento10 páginasGoma FinalHenrry CondeAún no hay calificaciones
- BIOGASDocumento19 páginasBIOGASRonald GamarraAún no hay calificaciones
- Estudio Sintético de Un Anabólico Androgénico, La Oxandrolona.Documento6 páginasEstudio Sintético de Un Anabólico Androgénico, La Oxandrolona.KariitoFerrerPertuzAún no hay calificaciones
- EmusionesDocumento11 páginasEmusionesangie ramirezAún no hay calificaciones
- Ficha Tecnica Bota Saga 2021Documento1 páginaFicha Tecnica Bota Saga 2021John Ever Quiroz Herrera100% (1)
- Cntaminacion Por Olores OfensivosDocumento29 páginasCntaminacion Por Olores OfensivosDaniela GomezAún no hay calificaciones
- Prueba Vida SaludableDocumento6 páginasPrueba Vida SaludableAirelav TN100% (1)
- No2 1Documento13 páginasNo2 1Ricardo HernandezAún no hay calificaciones
- Libro 1Documento23 páginasLibro 1Lopez R JaimeAún no hay calificaciones
- Guia Circulacion en Plantas y Hongos 6Documento6 páginasGuia Circulacion en Plantas y Hongos 6Angelita SuarezAún no hay calificaciones
- EugenolDocumento2 páginasEugenolKatherin ToapantaAún no hay calificaciones
- Upstream DownstreamDocumento7 páginasUpstream DownstreamCHANFLEAún no hay calificaciones
- Te. 10 Expresión Genética. Transcripción. Notas de Las DiapositivasDocumento8 páginasTe. 10 Expresión Genética. Transcripción. Notas de Las DiapositivasNayumi MavilaAún no hay calificaciones
- Practica Acidos Bases y Neutralizacion.Documento5 páginasPractica Acidos Bases y Neutralizacion.Areli SerranoAún no hay calificaciones
- Test Examen Completo BQDocumento114 páginasTest Examen Completo BQAlberto BalbinAún no hay calificaciones
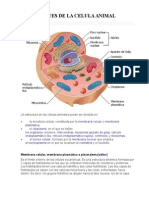
- Partes de La Celula AnimalDocumento7 páginasPartes de La Celula AnimalDanilo Daniel López PérezAún no hay calificaciones
- Determinación de Cenizas en AlimentosDocumento9 páginasDeterminación de Cenizas en AlimentostatianaAún no hay calificaciones
- 02 BacteriasDocumento41 páginas02 BacteriasIvette Pebes CAún no hay calificaciones
- Manual de Laboratorio Esc Ing. Ind. Quimica Organica Vilma Dianderas - Rossana JuarezDocumento58 páginasManual de Laboratorio Esc Ing. Ind. Quimica Organica Vilma Dianderas - Rossana JuarezHADY JAMILEE RAMOS MAMANIAún no hay calificaciones
- 7mo Informe Laboratorio de Quimica InroganicaDocumento13 páginas7mo Informe Laboratorio de Quimica InroganicaIsaac Farfan CondorAún no hay calificaciones
- Práctica 5 Optativa 3 PDFDocumento17 páginasPráctica 5 Optativa 3 PDFDiego moralesAún no hay calificaciones
- Curso Torres de DestilacionDocumento2 páginasCurso Torres de DestilacionandreaAún no hay calificaciones
- El Yodo Es Un Oxidante Medio y Permite Valorar Los TiosulfatosDocumento2 páginasEl Yodo Es Un Oxidante Medio y Permite Valorar Los TiosulfatosRosalia Pacheco CastroAún no hay calificaciones
- Ejemplos de Ácidos Carboxílicos SaturadosDocumento7 páginasEjemplos de Ácidos Carboxílicos SaturadosDenis Izquierdo HernandezAún no hay calificaciones
- Laboratorio 3. Química Orgánica IIDocumento3 páginasLaboratorio 3. Química Orgánica IISebastian VeraAún no hay calificaciones
- Aldehídos y CetonasDocumento21 páginasAldehídos y CetonasAndrews MenjivarAún no hay calificaciones
- Practicas QoiiDocumento39 páginasPracticas Qoiijuan islas100% (1)