También podría gustarte
- Temario Bioquímica - AngelDocumento30 páginasTemario Bioquímica - AngelMaría Hidalgo BorregoAún no hay calificaciones
- Teo - Diapositivas - Regulación de La Transcripción en EucariotasDocumento77 páginasTeo - Diapositivas - Regulación de La Transcripción en EucariotasMimi AcvredrAún no hay calificaciones
- Repaso de Biologia MolecularDocumento28 páginasRepaso de Biologia MolecularClaudia RamírezAún no hay calificaciones
- 10.regulación Génica en EucariotasDocumento20 páginas10.regulación Génica en EucariotasAgostina de MaríaAún no hay calificaciones
- Exposición CursoDocumento9 páginasExposición Cursojoha leivaAún no hay calificaciones
- Guia de VegetalDocumento3 páginasGuia de VegetalDaniela PadillaAún no hay calificaciones
- Capitulo 1. Embriología MédicaDocumento16 páginasCapitulo 1. Embriología MédicaYessenia MillaAún no hay calificaciones
- Transcripción Del ADNDocumento17 páginasTranscripción Del ADNdarwin plazaAún no hay calificaciones
- Ensayo Operones y ArniDocumento9 páginasEnsayo Operones y ArniAntonio Tenorio GonzalezAún no hay calificaciones
- Experimento de Beadle y TatumDocumento7 páginasExperimento de Beadle y TatumJuli SaplanaAún no hay calificaciones
- Tema 1 ING GDocumento3 páginasTema 1 ING GMARIA DEL CARMEN MOLINA HIGUERASAún no hay calificaciones
- Clase 1Documento53 páginasClase 1Mario MoralesAún no hay calificaciones
- Genes y CromosomasDocumento5 páginasGenes y Cromosomasjuan david torrealbaAún no hay calificaciones
- CÁP. 7 - Regulación de La Expresión GénicaDocumento5 páginasCÁP. 7 - Regulación de La Expresión GénicaJulia KatlenAún no hay calificaciones
- 4a. Procesamiento Del RNA en Procariotas y EucariotasDocumento34 páginas4a. Procesamiento Del RNA en Procariotas y EucariotasRaquel Pérez FernándezAún no hay calificaciones
- Silo - Tips - La Transcripcion en EucariotasDocumento25 páginasSilo - Tips - La Transcripcion en EucariotasDai DinamarcaAún no hay calificaciones
- Capitulo3 - Capitulo 4Documento10 páginasCapitulo3 - Capitulo 4Laura UzumakiAún no hay calificaciones
- La Célula VegetalDocumento9 páginasLa Célula VegetalNatalia EspinarAún no hay calificaciones
- DOGMA CENTRAL, Transcripción y TraducciónDocumento17 páginasDOGMA CENTRAL, Transcripción y TraducciónK. Lizeth G.CAún no hay calificaciones
- Cuestionario Cuestionario Acidos NucleicosDocumento3 páginasCuestionario Cuestionario Acidos NucleicosNandoFernandoAún no hay calificaciones
- La Triple Hélice en Terapia Génica - Lía Cristina Upegui-GonzálezDocumento14 páginasLa Triple Hélice en Terapia Génica - Lía Cristina Upegui-Gonzálezsanti333100% (1)
- Control de La Expresión GenéticaDocumento8 páginasControl de La Expresión GenéticaCamila MonciónAún no hay calificaciones
- Resumen de Regulacion A Nivel DNADocumento11 páginasResumen de Regulacion A Nivel DNALuis Antonio Perez AvitiaAún no hay calificaciones
- Genética. Eje 5. Transcripción y TraducciónDocumento26 páginasGenética. Eje 5. Transcripción y TraducciónCecilia Quiroga De DavilaAún no hay calificaciones
- Rejunte de 2 Praciales BIOQUIM IIDocumento6 páginasRejunte de 2 Praciales BIOQUIM IIMaia CucchianiAún no hay calificaciones
- Preguntas Regulacion de La Expresion GenicaDocumento3 páginasPreguntas Regulacion de La Expresion GenicaHelen Romero100% (3)
- Examen 1 de Biotecnología - Sergio de Jesús Jaime RodríguezDocumento3 páginasExamen 1 de Biotecnología - Sergio de Jesús Jaime RodríguezSergio de Jesús Jaime RodríguezAún no hay calificaciones
- Genetica Clase 2Documento6 páginasGenetica Clase 2Paula DiegoAún no hay calificaciones
- Regulación de La Expresión GénicaDocumento34 páginasRegulación de La Expresión GénicamiriammoreyAún no hay calificaciones
- Taller Expresión GénicaDocumento10 páginasTaller Expresión GénicaEdith GuerreroAún no hay calificaciones
- Elvia Isabel Guacaneme Perez, Taller 5Documento4 páginasElvia Isabel Guacaneme Perez, Taller 5Elvia Isabel GUACANEME PEREZAún no hay calificaciones
- Actividad 3, GeneticaDocumento7 páginasActividad 3, GeneticaSheccid MedinaAún no hay calificaciones
- Regulación TranscripcionalDocumento31 páginasRegulación TranscripcionalFermín Armando Ledezma VelázquezAún no hay calificaciones
- Resumen de Embriología Regulacion MolecularDocumento5 páginasResumen de Embriología Regulacion MolecularJeisson Jauregui100% (1)
- Operón BacterianoDocumento2 páginasOperón BacterianoAdrianAlexanderChangalomboTravezAún no hay calificaciones
- Las Hormonas Pueden Influir Sobre Efectos Biológicos Específicos Al Modular La TranscripciónDocumento6 páginasLas Hormonas Pueden Influir Sobre Efectos Biológicos Específicos Al Modular La TranscripciónHéctor ZaldívarAún no hay calificaciones
- Parcial Unidad 2 - Genética GeneralDocumento7 páginasParcial Unidad 2 - Genética GeneralMÓNICA VIVIANA GÓMEZ VARGASAún no hay calificaciones
- TelomerosDocumento9 páginasTelomerosCristian Regis Torres FriasAún no hay calificaciones
- Introducción A La Regulación y Señalización MolecularDocumento41 páginasIntroducción A La Regulación y Señalización MolecularMarina Itzel Castillo Ambriz50% (2)
- Los Estimuladores y Silenciadores Elementos de RegulaciónDocumento3 páginasLos Estimuladores y Silenciadores Elementos de RegulaciónDIEGO MAURICIO SALAZAR ZAMBRANOAún no hay calificaciones
- La Regulación de La Traducción de ProteínasDocumento4 páginasLa Regulación de La Traducción de ProteínasLUISA DANIELA CACERES RUBIOAún no hay calificaciones
- TALLER DE PREGUNTAS Biologia Molecular 3Documento5 páginasTALLER DE PREGUNTAS Biologia Molecular 3Diego RodriguezAún no hay calificaciones
- ADN SuperenrolladoDocumento203 páginasADN Superenrolladofrancisco lozano gomezAún no hay calificaciones
- Clase 8.1 EpigénicaDocumento7 páginasClase 8.1 EpigénicaClaudia BAún no hay calificaciones
- Epigenética AndrésDocumento30 páginasEpigenética AndrésAlberto BeskowAún no hay calificaciones
- ARN EUCARIOTA EspañolDocumento14 páginasARN EUCARIOTA EspañolHector SustaydAún no hay calificaciones
- Taller 6. Regulación de La Expresión en EucariotasDocumento4 páginasTaller 6. Regulación de La Expresión en EucariotasPaula NastarAún no hay calificaciones
- Bitacora-Porta Gen Mol U3 EmmanuelDocumento37 páginasBitacora-Porta Gen Mol U3 EmmanuelEmmanuel GutiérrezAún no hay calificaciones
- Biología Molecular Avanzada IDocumento20 páginasBiología Molecular Avanzada ICristobal GallardoAún no hay calificaciones
- POLYOMAVIRUSES 2 EsDocumento11 páginasPOLYOMAVIRUSES 2 EsMICHELL CAROLINA FERNANDEZ PAEZAún no hay calificaciones
- La Función de GenBIOMAGNETDocumento28 páginasLa Función de GenBIOMAGNETtito zambranoAún no hay calificaciones
- Clase 2 Ohler2009Documento12 páginasClase 2 Ohler2009Natalia Andrea Nuñez ToledoAún no hay calificaciones
- Los Factores Generales de Transcripción Del ARN Polimerasa IIDocumento21 páginasLos Factores Generales de Transcripción Del ARN Polimerasa IIcsos csosAún no hay calificaciones
- MonografiaDocumento4 páginasMonografiaJose Juan Torres MartinezAún no hay calificaciones
- Wuolah Free TEMA 10 Regulacion de La Exresion Genica en Eucariotas Gulag FreeDocumento9 páginasWuolah Free TEMA 10 Regulacion de La Exresion Genica en Eucariotas Gulag FreeMarta Elisa Sánchez GallardoAún no hay calificaciones
- Transcripción y TraducciónDocumento6 páginasTranscripción y TraducciónJose Luis VallejosAún no hay calificaciones
- Biología Molecular Aplicada A Alimentos - Tema 1Documento5 páginasBiología Molecular Aplicada A Alimentos - Tema 1Florina Iulia BuraAún no hay calificaciones
- Qué Es y Que Función Tiene Un Factor de TranscripciónDocumento3 páginasQué Es y Que Función Tiene Un Factor de TranscripciónMaria Elizabeth GaliciaAún no hay calificaciones
- Más allá del ADN: La Revolución Epigenética: Desde Mecanismos Celulares hasta Factores Ambientales: Cómo la Epigenética Moldea Nuestro Destino Biológico y las Implicaciones para la Salud, el Comportamiento y el Futuro de la InvestigaciónDe EverandMás allá del ADN: La Revolución Epigenética: Desde Mecanismos Celulares hasta Factores Ambientales: Cómo la Epigenética Moldea Nuestro Destino Biológico y las Implicaciones para la Salud, el Comportamiento y el Futuro de la InvestigaciónAún no hay calificaciones
- Planos Del Servicio Farmaceutico y Utillaje de Cada AreaDocumento15 páginasPlanos Del Servicio Farmaceutico y Utillaje de Cada Arealiz920779% (14)
- ENSAYODocumento1 páginaENSAYOliz9207Aún no hay calificaciones
- Acción Del Sol en La Piel y El CabelloDocumento10 páginasAcción Del Sol en La Piel y El Cabelloliz9207Aún no hay calificaciones
- Involución CutáneaDocumento13 páginasInvolución Cutánealiz9207Aún no hay calificaciones
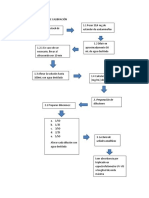
- Flujograma de Curva de CalibraciónDocumento1 páginaFlujograma de Curva de Calibraciónliz9207Aún no hay calificaciones
- ENSAYODocumento1 páginaENSAYOliz9207Aún no hay calificaciones
- Ejemplo 13 Parte ADocumento2 páginasEjemplo 13 Parte Aliz9207Aún no hay calificaciones
- Articulo CompDocumento11 páginasArticulo Compliz9207Aún no hay calificaciones
- Las 9estrellasDocumento3 páginasLas 9estrellasliz9207100% (1)
- GraficasDocumento9 páginasGraficasliz9207Aún no hay calificaciones
- Eum InformeDocumento17 páginasEum Informeliz9207Aún no hay calificaciones
- Protein AsDocumento25 páginasProtein Asliz9207Aún no hay calificaciones
- Articulo Quimica ComputacionalDocumento6 páginasArticulo Quimica Computacionalliz9207Aún no hay calificaciones
- Solventes PolaresDocumento1 páginaSolventes Polaresliz9207Aún no hay calificaciones
- Apendice 4Documento4 páginasApendice 4liz9207Aún no hay calificaciones
- Otros Fármacos AgonistasDocumento6 páginasOtros Fármacos Agonistasliz9207Aún no hay calificaciones
- Otros Fármacos AgonistasDocumento6 páginasOtros Fármacos Agonistasliz9207Aún no hay calificaciones
- Teoras Del Currculo y Concepciones CurricularesDocumento22 páginasTeoras Del Currculo y Concepciones CurricularesmilderavilaAún no hay calificaciones
- Educacion para La Salud Folletooo PDFDocumento2 páginasEducacion para La Salud Folletooo PDFliz9207Aún no hay calificaciones
- Modelos Experimentales para Evaluar de La PermeabilidadDocumento10 páginasModelos Experimentales para Evaluar de La Permeabilidadliz9207Aún no hay calificaciones
- Apendice 4Documento4 páginasApendice 4liz9207Aún no hay calificaciones
- FarmacéuticaDocumento8 páginasFarmacéuticaliz9207Aún no hay calificaciones
- Resumen TLRDocumento5 páginasResumen TLRliz9207Aún no hay calificaciones
- Eliminación de FármacosDocumento12 páginasEliminación de Fármacosliz9207Aún no hay calificaciones
- Reacción de CaCO3 Con Ácido ClorhídricoDocumento6 páginasReacción de CaCO3 Con Ácido Clorhídricoliz9207Aún no hay calificaciones
- Practica 7 Determinacion de ColesterolDocumento16 páginasPractica 7 Determinacion de Colesterolliz9207100% (1)
- Informe de Laboratorio LizmethDocumento6 páginasInforme de Laboratorio Lizmethliz9207Aún no hay calificaciones
- FenolesDocumento6 páginasFenolesliz9207Aún no hay calificaciones
- Practica 5 Determinacion de EnzimaDocumento16 páginasPractica 5 Determinacion de Enzimaliz92070% (1)
- Clase Clasificación de Los VirusDocumento10 páginasClase Clasificación de Los VirusMaría Fernanda Mojica CastilloAún no hay calificaciones
- PH y Concentrecion EnzimaticaDocumento3 páginasPH y Concentrecion EnzimaticaSaul SalasAún no hay calificaciones
- Clase 3Documento53 páginasClase 3Rober ParedesAún no hay calificaciones
- Práctica Ii - Guía - Proteínas IDocumento5 páginasPráctica Ii - Guía - Proteínas IMarjorie Lizbeth P.100% (1)
- Cap 42Documento11 páginasCap 42CARE LESSAún no hay calificaciones
- Genética, Historia y FuturoDocumento3 páginasGenética, Historia y FuturoIsabela Giraldo VillegasAún no hay calificaciones
- Practica7 Muñoz SiguenzaDocumento11 páginasPractica7 Muñoz SiguenzaSasha Valeria Siguenza RoblesAún no hay calificaciones
- Guía#3 Noveno 2023Documento6 páginasGuía#3 Noveno 2023Sofia Serrano GalindoAún no hay calificaciones
- El Genoma HumanoDocumento5 páginasEl Genoma HumanoDORA ALICIA ALVA LAVALLE100% (1)
- Práctica 8 PDFDocumento4 páginasPráctica 8 PDFJaneth RodriguezAún no hay calificaciones
- INHIBIDORESDocumento3 páginasINHIBIDORESLESLIE EVELIN LOMA JIMENEZAún no hay calificaciones
- Taller de SinapsisDocumento4 páginasTaller de Sinapsismatacurie100% (1)
- Guía La Celula Grado 11-2021Documento7 páginasGuía La Celula Grado 11-2021Karla F. RomeroAún no hay calificaciones
- Replicación de ADN BAmbDocumento13 páginasReplicación de ADN BAmbMaldonado López GabrielaAún no hay calificaciones
- Oncogénesis PDFDocumento19 páginasOncogénesis PDFlilianapradaAún no hay calificaciones
- Capitulo 18Documento11 páginasCapitulo 18Carlos HomeAún no hay calificaciones
- NucleoplasmaDocumento2 páginasNucleoplasmaCelular Dos MilAún no hay calificaciones
- Tarea 1. Abeso MangueDocumento10 páginasTarea 1. Abeso ManguePedro Ernesto Abeso MangueAún no hay calificaciones
- 01 BiomembranasDocumento7 páginas01 BiomembranasKaren RamosAún no hay calificaciones
- Aminoácidos - PéptidosDocumento27 páginasAminoácidos - Péptidosjhonny pascualAún no hay calificaciones
- Nucleotidos y NucleoprotreinasDocumento38 páginasNucleotidos y NucleoprotreinasNazareth LucianaAún no hay calificaciones
- Gel Basado en PapainaDocumento15 páginasGel Basado en PapainaDanny DanAún no hay calificaciones
- Tecnologia-Del-Adn-RecombinanteDocumento16 páginasTecnologia-Del-Adn-RecombinanteGisela Anampa MonzonAún no hay calificaciones
- 01 Matriz CitoplasmaticaDocumento44 páginas01 Matriz CitoplasmaticaAbimael Choqque TorresAún no hay calificaciones
- Guía de Actividades y Rúbrica de Evaluación - Tarea 1 - BiomoléculasDocumento11 páginasGuía de Actividades y Rúbrica de Evaluación - Tarea 1 - BiomoléculasLaudith OrtegaAún no hay calificaciones
- Los Ácidos Nuclicos y Sus ProcesosDocumento23 páginasLos Ácidos Nuclicos y Sus Procesosluz claraAún no hay calificaciones
- Mapa Conceptual, Generalidades de La EmbriologiaDocumento2 páginasMapa Conceptual, Generalidades de La Embriologiajuan CeTruAún no hay calificaciones
- Enzimas PDFDocumento53 páginasEnzimas PDFNelson BeltranAún no hay calificaciones
- Met 3 - VECTORES VIRALES PARA LA TRANSDUCCIÓN CELULARDocumento1 páginaMet 3 - VECTORES VIRALES PARA LA TRANSDUCCIÓN CELULARPatricia GarcíaAún no hay calificaciones
- Mapa Conceptual Sobre Clasificación de Las ProteínasDocumento1 páginaMapa Conceptual Sobre Clasificación de Las ProteínasDulce María Santos MenjivarAún no hay calificaciones