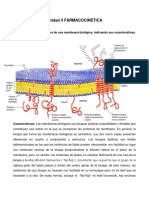
También podría gustarte
- Diferencias Hombre Mujer: Descubre Los Últimos Hallazgos Científicos Sobre Las Diferencias Entre Mujeres Y HombresDe EverandDiferencias Hombre Mujer: Descubre Los Últimos Hallazgos Científicos Sobre Las Diferencias Entre Mujeres Y HombresAún no hay calificaciones
- Examen de Farmacología bloques 1 y 2Documento78 páginasExamen de Farmacología bloques 1 y 2Flavia Chavez Almaraz100% (3)
- Informe N°3. Analisis Microbiologico de Alimentos.Documento7 páginasInforme N°3. Analisis Microbiologico de Alimentos.JAVIER ESTEBAN ARGOTY ORTEGONAún no hay calificaciones
- Biologia 5º II VolumenDocumento32 páginasBiologia 5º II VolumenDionne BeckAún no hay calificaciones
- Cromatina X de La Mucosa BucalDocumento4 páginasCromatina X de La Mucosa Bucal.:("*"BLacK BuLLeT"*"):.Aún no hay calificaciones
- Vertido en Placa y Extensión en Placa ComparacionDocumento1 páginaVertido en Placa y Extensión en Placa ComparacionJuan Volta Bellamy100% (1)
- Sindrome de TurnerDocumento18 páginasSindrome de TurnerNicole Diaz100% (2)
- MutacionDocumento4 páginasMutacionmaria joseAún no hay calificaciones
- Inactivación del Cromosoma X enDocumento4 páginasInactivación del Cromosoma X enAlejandra OvalleAún no hay calificaciones
- Cromosoma X inactivo y enfermedadesDocumento12 páginasCromosoma X inactivo y enfermedadesTatiana BerrocalAún no hay calificaciones
- Cromatina XDocumento19 páginasCromatina XvbdgddgfdfgAún no hay calificaciones
- Corpusculos de Barr en Células de La Mucosa OralDocumento10 páginasCorpusculos de Barr en Células de La Mucosa OralMiguel ParejoAún no hay calificaciones
- Capitulo 7 KlugDocumento10 páginasCapitulo 7 KlugIngrid Anaya MoralesAún no hay calificaciones
- Cromosomas X: Lyonización y corpúsculos de BarrDocumento5 páginasCromosomas X: Lyonización y corpúsculos de BarrVale HAún no hay calificaciones
- Seminario Genética Caso de Gatos CalicóDocumento23 páginasSeminario Genética Caso de Gatos CalicóRodrigo AntilAún no hay calificaciones
- Cuestionario Practica 2Documento2 páginasCuestionario Practica 2Courtney PetersonAún no hay calificaciones
- Cromatina SexualDocumento19 páginasCromatina SexualTania GonzalezAún no hay calificaciones
- Corpusculo BarrDocumento5 páginasCorpusculo BarrFrailejon Ernesto PerezAún no hay calificaciones
- Cromosoma X Inactivación Compensación Dosis GénicaDocumento18 páginasCromosoma X Inactivación Compensación Dosis GénicaPercy Ureta SierraAún no hay calificaciones
- Guias Aberraciones Cr. SexualesDocumento16 páginasGuias Aberraciones Cr. Sexualeseze pedrozoAún no hay calificaciones
- Principio de LyonDocumento15 páginasPrincipio de Lyonalzheimer382Aún no hay calificaciones
- Cromatina SexualDocumento3 páginasCromatina SexualValeria DelgadilloAún no hay calificaciones
- Cromosoma XDocumento8 páginasCromosoma XMarita ToctoAún no hay calificaciones
- Inactivacion Del Cromosoma XDocumento8 páginasInactivacion Del Cromosoma XAldair MedinaAún no hay calificaciones
- Inactivacion Cromosoma X Las MejoresDocumento20 páginasInactivacion Cromosoma X Las MejoresAlexandra Saona MarínAún no hay calificaciones
- Genetica - Cromosomas X y YDocumento50 páginasGenetica - Cromosomas X y YMichael QuinteroAún no hay calificaciones
- Determinación del sexo en células epiteliales mediante la identificación de la cromatina XDocumento6 páginasDeterminación del sexo en células epiteliales mediante la identificación de la cromatina XSofia MartínezAún no hay calificaciones
- Identificación de la cromatina sexual femeninaDocumento6 páginasIdentificación de la cromatina sexual femeninaTatii Vega Cañate100% (1)
- Inactivacion XDocumento34 páginasInactivacion XLizbeth DavilaAún no hay calificaciones
- Cromosomas sexuales y aneuploidíasDocumento7 páginasCromosomas sexuales y aneuploidíasRaymundo SaldañaAún no hay calificaciones
- Informe Corpúsculo de BarrDocumento9 páginasInforme Corpúsculo de BarrJorge Blanquiceth100% (1)
- Cuerpos de Barr InformeDocumento9 páginasCuerpos de Barr Informejose coronado gonzalezAún no hay calificaciones
- Tarea 2 - 27 - JessicaElizabethPérezLimaDocumento2 páginasTarea 2 - 27 - JessicaElizabethPérezLimaElizabeth Pérez LimaAún no hay calificaciones
- Inactivación Del Cromosoma X y DiabetesDocumento33 páginasInactivación Del Cromosoma X y Diabetessindy garciaAún no hay calificaciones
- Inmuno Practica 1 y 2Documento6 páginasInmuno Practica 1 y 2Emmanuel Rodriguez MontañezAún no hay calificaciones
- Cromatina X y diagnóstico toxoplasmosisDocumento5 páginasCromatina X y diagnóstico toxoplasmosisNati Fernandez100% (1)
- Tema 4, 5 Genética MédicaDocumento17 páginasTema 4, 5 Genética MédicaEme PintoAún no hay calificaciones
- Síndrome de Turner2016 - 5 - 30P15 - 30 - 27Documento2 páginasSíndrome de Turner2016 - 5 - 30P15 - 30 - 27Imanol TrejoAún no hay calificaciones
- LA GENETICA NO MENDELIANA (Parte2)Documento2 páginasLA GENETICA NO MENDELIANA (Parte2)YULIANY ANDREA ARCINIEGAS ISCALAAún no hay calificaciones
- Cromatina SexualDocumento37 páginasCromatina SexualPenélope PinedaAún no hay calificaciones
- PRÁCTICASDocumento13 páginasPRÁCTICASEpicMilfTime100% (4)
- Comprension LectoraDocumento4 páginasComprension LectoraFrancisco AravenaAún no hay calificaciones
- Tema 14 Herencia Ligada Al SexoDocumento33 páginasTema 14 Herencia Ligada Al Sexocrissierrarodas1Aún no hay calificaciones
- Rasgos Ligados Al SexoDocumento7 páginasRasgos Ligados Al SexoYang LightAún no hay calificaciones
- Patrones herencia MendelDocumento10 páginasPatrones herencia MendelMari Angeles VargasAún no hay calificaciones
- Modos de Herencia Ligados Al Sexo1Documento8 páginasModos de Herencia Ligados Al Sexo1Bryan JacomeAún no hay calificaciones
- Cromosomas Sexuales, Determinacion Genetica Del SexoDocumento4 páginasCromosomas Sexuales, Determinacion Genetica Del SexoJime GrossoAún no hay calificaciones
- Corpusculo de BarrDocumento8 páginasCorpusculo de BarrJesus Alberto Castro BohorquezAún no hay calificaciones
- Sindrome de KlinefelterDocumento55 páginasSindrome de Klinefelterabdelbrasileiro2001Aún no hay calificaciones
- Fisiopatologia, Unidad 1Documento38 páginasFisiopatologia, Unidad 1Diego PintoAún no hay calificaciones
- Informe Cromatina Sexual.Documento7 páginasInforme Cromatina Sexual.Edgar Andrade Zambrano50% (4)
- Cromatina X o Corpúsculo de BarrDocumento11 páginasCromatina X o Corpúsculo de BarrChe Luiz T Castañeda0% (1)
- GM - Guía de Estudio #4Documento12 páginasGM - Guía de Estudio #4mczanaboni100% (2)
- Inactivacion Del Cromosoma XDocumento6 páginasInactivacion Del Cromosoma XiaobregonAún no hay calificaciones
- Taller Herencia Del Sexo y Ligamiento y Entrecuzamiento Arzuaga - Cantillo - Galindo - ZapataDocumento19 páginasTaller Herencia Del Sexo y Ligamiento y Entrecuzamiento Arzuaga - Cantillo - Galindo - ZapataMayllin HortaAún no hay calificaciones
- Trastornos Del Desarrollo SexualDocumento4 páginasTrastornos Del Desarrollo SexualCarmen BustillosAún no hay calificaciones
- Sindrome de KlinefelterDocumento17 páginasSindrome de KlinefelterMiguel MarroquínAún no hay calificaciones
- GENETICA HemofiliaDocumento30 páginasGENETICA Hemofiliajose duarteAún no hay calificaciones
- Genética - Herencia ligada al sexo y determinación sexualDocumento13 páginasGenética - Herencia ligada al sexo y determinación sexualSaraAún no hay calificaciones
- Tema 9Documento6 páginasTema 9Elisa Barreiro DocioAún no hay calificaciones
- Inactivacion Del Cromosoma X Hipotesis de LyonDocumento7 páginasInactivacion Del Cromosoma X Hipotesis de LyonJorge MoscosoAún no hay calificaciones
- Cuerpo de BarrDocumento4 páginasCuerpo de BarrBORIS MASTERAún no hay calificaciones
- Benjamin A Pierce FUNDAMENTOS DE GENETICDocumento3 páginasBenjamin A Pierce FUNDAMENTOS DE GENETICalanjosue755Aún no hay calificaciones
- Tare 1.2 NURS 1050Documento8 páginasTare 1.2 NURS 1050Delvis RodriguezAún no hay calificaciones
- NDP Peset-Isasa DefDocumento2 páginasNDP Peset-Isasa DefmonicaAún no hay calificaciones
- Tarea Farma. Unidad 4Documento30 páginasTarea Farma. Unidad 4Rachel MedAún no hay calificaciones
- 2 Biología SextoDocumento6 páginas2 Biología Sextoparkour parkourAún no hay calificaciones
- Conceptos Basicos de Señalizacion Molecular Del DesarrolloDocumento6 páginasConceptos Basicos de Señalizacion Molecular Del DesarrolloItzel HernándezAún no hay calificaciones
- Todas Las Células Provienen de Otras CélulasDocumento1 páginaTodas Las Células Provienen de Otras CélulasMichael Joseph EsquivelAún no hay calificaciones
- Clase 16 Concepto de Embriogenesis e HistogenesisDocumento16 páginasClase 16 Concepto de Embriogenesis e HistogenesisJuan Pablo LempAún no hay calificaciones
- Unid III. Proteinas 201920 - MovilidadDocumento31 páginasUnid III. Proteinas 201920 - MovilidadSEBASTIAN ALONSO ORTIZ MATTAAún no hay calificaciones
- Resumen Del Código Genético BilogiaDocumento3 páginasResumen Del Código Genético BilogiaWilfran OrtizAún no hay calificaciones
- Guia de Actividades y Rúbrica de Evaluación - Unidad 3 - Tarea 4 - AnalizarDocumento9 páginasGuia de Actividades y Rúbrica de Evaluación - Unidad 3 - Tarea 4 - AnalizarDiana Marcela Ahumada SánchezAún no hay calificaciones
- Practica #9Documento6 páginasPractica #9Ysabel Borda DonairesAún no hay calificaciones
- Cromosomas Sexuales, Determinacion Genetica Del SexoDocumento4 páginasCromosomas Sexuales, Determinacion Genetica Del SexoJime GrossoAún no hay calificaciones
- Silabo de BioquímicaDocumento6 páginasSilabo de BioquímicaDilan RuedaAún no hay calificaciones
- Agente Etiológico VihDocumento4 páginasAgente Etiológico VihKaren VeraAún no hay calificaciones
- Metabolismo Despúes de La Cosecha. IND. I FRUTAS Y HORTALIZAS - 06 NOVIEMBRE 2020Documento4 páginasMetabolismo Despúes de La Cosecha. IND. I FRUTAS Y HORTALIZAS - 06 NOVIEMBRE 2020Hjalmar LinoAún no hay calificaciones
- Guía Control Ciclo Celular y CancerDocumento5 páginasGuía Control Ciclo Celular y CancerpepepepinoAún no hay calificaciones
- Clase 10Documento18 páginasClase 10Nicole TurraAún no hay calificaciones
- 20 Semanas de MicrobiologiaDocumento33 páginas20 Semanas de MicrobiologiaAlejandro YañezAún no hay calificaciones
- 2006 Libro Citogenetica de PecesDocumento204 páginas2006 Libro Citogenetica de PecesJonathanSanchezLopezAún no hay calificaciones
- EVOLUCIÓNDocumento80 páginasEVOLUCIÓNxSetterAún no hay calificaciones
- FRASES INDIGENAS Y GENÉTICADocumento1 páginaFRASES INDIGENAS Y GENÉTICAArturo CidAún no hay calificaciones
- Examen GeneticaDocumento2 páginasExamen GeneticaSonia CalvoAún no hay calificaciones
- Diversidad y Biodiversidad Autor: R. Margalef: Lima, PerúDocumento2 páginasDiversidad y Biodiversidad Autor: R. Margalef: Lima, PerúCarlosAún no hay calificaciones
- Ramas de La ZoologiaDocumento3 páginasRamas de La Zoologiaisrael camposAún no hay calificaciones
- PCR: Reacción en Cadena de la PolimerasaDocumento42 páginasPCR: Reacción en Cadena de la PolimerasaDiego SevillaAún no hay calificaciones