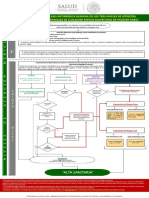
También podría gustarte
- Monografia La TuberculosisDocumento20 páginasMonografia La TuberculosisHugo Richard Flores Contreras83% (23)
- Ut3. Aplicación de Técnicas de Comunicación y Apoyo PsicológicoDocumento21 páginasUt3. Aplicación de Técnicas de Comunicación y Apoyo PsicológicoPilar MonteroAún no hay calificaciones
- GPC Tuberculosis PulmonarDocumento10 páginasGPC Tuberculosis PulmonarOvejita GaleanaAún no hay calificaciones
- Examen de Recuperación Formacion Civica y Etica IiDocumento10 páginasExamen de Recuperación Formacion Civica y Etica IiLino Cambrón100% (4)
- Guías de Practicas Clinicas EstomatológicasDocumento73 páginasGuías de Practicas Clinicas EstomatológicasRip Vera67% (9)
- Tuberculosis: Natural History, Microbiology, and PathogenesisDocumento24 páginasTuberculosis: Natural History, Microbiology, and PathogenesisIdaly CoronaAún no hay calificaciones
- Tuberculosis PulmonarDocumento12 páginasTuberculosis PulmonarVictor MonsalveAún no hay calificaciones
- Tuberculosis BacteriologiaDocumento15 páginasTuberculosis BacteriologiaVillagomez López Alejandra AracelyAún no hay calificaciones
- Microbiología y Patogénesis de La TuberculosisDocumento17 páginasMicrobiología y Patogénesis de La Tuberculosiscd1805Aún no hay calificaciones
- TUBERCULOSISDocumento35 páginasTUBERCULOSISSteven Catunta ChoqueAún no hay calificaciones
- TBC - DiaspositivasDocumento50 páginasTBC - DiaspositivasCarlos Barreto CervantesAún no hay calificaciones
- TB Urogenital Wilson IdrovoDocumento24 páginasTB Urogenital Wilson IdrovobryanAún no hay calificaciones
- La TuberculosisDocumento25 páginasLa TuberculosisLINAAún no hay calificaciones
- Introducción Ala TuberculosisDocumento26 páginasIntroducción Ala TuberculosisAlex Franco Leonardo Shahuano0% (1)
- Infecciones Especificas Del Tracto UrogenitalDocumento32 páginasInfecciones Especificas Del Tracto UrogenitalAlexis Garcia CasusolAún no hay calificaciones
- Trabajo NefroDocumento23 páginasTrabajo NefroGlaudiovane JúniorAún no hay calificaciones
- Triptico TBCDocumento12 páginasTriptico TBCInfanteria de MarinaAún no hay calificaciones
- Etiopatogenia de La TBCDocumento6 páginasEtiopatogenia de La TBCAmelia BarrantesAún no hay calificaciones
- 1 TuberculosisDocumento5 páginas1 TuberculosisDorian Victor Encinas VargasAún no hay calificaciones
- Epidemiología de La TuberculosisDocumento14 páginasEpidemiología de La TuberculosisEDGAR ROLANDO MACZ USHERAún no hay calificaciones
- Epidemiología de La TuberculosisDocumento16 páginasEpidemiología de La TuberculosislissitaprzAún no hay calificaciones
- TBC1Documento6 páginasTBC1Natalia QuijadaAún no hay calificaciones
- Investigación - TuberculosisDocumento8 páginasInvestigación - TuberculosisSofia Bustos GaldoAún no hay calificaciones
- Guia - TBC SNC (Minsa)Documento33 páginasGuia - TBC SNC (Minsa)Alfonso A. Ortiz PatiñoAún no hay calificaciones
- Proyecto de TuberculosisDocumento22 páginasProyecto de TuberculosisDelfy ChoroAún no hay calificaciones
- A Escala MundialDocumento4 páginasA Escala MundialNOELIA JANET FLORES DURANDAún no hay calificaciones
- PATOLOGIA TUBERCULOSIS PULMONAR 8vo SEMESTREDocumento44 páginasPATOLOGIA TUBERCULOSIS PULMONAR 8vo SEMESTREDELMIRA ZAMBRANA VEIZAGAAún no hay calificaciones
- 7A Eq 1 Tuberculosis Extrapulmonar y Las Formas de Presentación PDFDocumento33 páginas7A Eq 1 Tuberculosis Extrapulmonar y Las Formas de Presentación PDFCristian MerchánAún no hay calificaciones
- Tuberculosis IIDocumento22 páginasTuberculosis IIVictor Daniel Pelagio QuintanaAún no hay calificaciones
- La Tuberculosis Constituye Un Paradigma de La Interacción de Un Agente Exógeno y La Respuesta Inmunitaria Del HuéspedDocumento1 páginaLa Tuberculosis Constituye Un Paradigma de La Interacción de Un Agente Exógeno y La Respuesta Inmunitaria Del HuéspedEstefanýa SasintuñaAún no hay calificaciones
- Tuberculosis Pier Expo 04-05-23Documento19 páginasTuberculosis Pier Expo 04-05-23Pier Jhoani Ñañez LlanosAún no hay calificaciones
- Patologia EspecialDocumento11 páginasPatologia EspecialVictoor JuaanAún no hay calificaciones
- TBCDocumento17 páginasTBCLid Lapa AlvarezAún no hay calificaciones
- Guia de Practica Clinica - TBC Sistema NerviosoDocumento38 páginasGuia de Practica Clinica - TBC Sistema NerviosoNalemi JTAún no hay calificaciones
- Monografia Tuberculosis. 2Documento28 páginasMonografia Tuberculosis. 2breidyAún no hay calificaciones
- Apa de AnatomiaDocumento15 páginasApa de AnatomiaROCIEL BRISA BULEJE PACURIAún no hay calificaciones
- T7 Tuberculosis IDocumento8 páginasT7 Tuberculosis IHematologia InclanAún no hay calificaciones
- Practica 13. Tuberculosis 2Documento11 páginasPractica 13. Tuberculosis 2Maria AldapeAún no hay calificaciones
- Tuberculosis Udabol2023Documento11 páginasTuberculosis Udabol2023rosmery supa floresAún no hay calificaciones
- TUBERCULOSISDocumento9 páginasTUBERCULOSISBerth GuachałłaAún no hay calificaciones
- TBC Dra LeccaDocumento45 páginasTBC Dra LeccaluceropurizacafiestasAún no hay calificaciones
- TUBERCULOSISresumenDocumento17 páginasTUBERCULOSISresumenMery MogollonAún no hay calificaciones
- Expo de Seminaro 2Documento20 páginasExpo de Seminaro 2Melissa ReyesAún no hay calificaciones
- Tuberculosis y EpilepsiaDocumento17 páginasTuberculosis y EpilepsiaJoseAngel19Aún no hay calificaciones
- TUBERCULOSISDocumento64 páginasTUBERCULOSISdayanaAún no hay calificaciones
- TBC - POWER POINT FilooooDocumento16 páginasTBC - POWER POINT FilooooEDFloresAún no hay calificaciones
- TuberculosisDocumento7 páginasTuberculosisjulissaAún no hay calificaciones
- Httpsbancos - Salud.gob - Arsitesdefaultfiles2018-100000000049cnt-Guia de Diagnostico Tratamiento y Prevencion de La TubercuDocumento70 páginasHttpsbancos - Salud.gob - Arsitesdefaultfiles2018-100000000049cnt-Guia de Diagnostico Tratamiento y Prevencion de La TubercucarolinagerezzAún no hay calificaciones
- Uber Culo SisDocumento14 páginasUber Culo SisFabricioVilcaAún no hay calificaciones
- TuberculosisDocumento15 páginasTuberculosisROLANDO PINTOAún no hay calificaciones
- Ensayo de La TuberculosisDocumento2 páginasEnsayo de La Tuberculosisraul vedia100% (2)
- TUBERCULOSISDocumento23 páginasTUBERCULOSISdanielaAún no hay calificaciones
- Seminario Adulto I FinallDocumento9 páginasSeminario Adulto I FinallZELADA CASAS MARCO ANTONIOAún no hay calificaciones
- 2021 Tuberculosis LemaDocumento6 páginas2021 Tuberculosis Lemajuany gonzalezAún no hay calificaciones
- Etiopatogenia y Fisiopatología de La TuberculosisDocumento9 páginasEtiopatogenia y Fisiopatología de La Tuberculosislulita1202Aún no hay calificaciones
- Tuberculosis Renal (Completo)Documento35 páginasTuberculosis Renal (Completo)Aivy Abanto MoyaAún no hay calificaciones
- Tuberculosis Caso ClinicoDocumento39 páginasTuberculosis Caso ClinicoYlsa Patiño0% (1)
- TuberculosisDocumento9 páginasTuberculosisgianela garciaAún no hay calificaciones
- TuberculosisDocumento8 páginasTuberculosisapi-3814371100% (4)
- Tuberculosis 2018Documento18 páginasTuberculosis 2018Margori Stefani Damian Becerra100% (1)
- Ensayo de TuberculosisDocumento5 páginasEnsayo de TuberculosisOliverDurazo50% (2)
- Infección Tuberculosa Latente, la base del icebergDe EverandInfección Tuberculosa Latente, la base del icebergAún no hay calificaciones
- La pandemia del covid-19. Efectos sobre la saludDe EverandLa pandemia del covid-19. Efectos sobre la saludCalificación: 5 de 5 estrellas5/5 (1)
- The Natural History and Clinical Features of HIV Infection in Adults and Adolescents - UpToDateDocumento40 páginasThe Natural History and Clinical Features of HIV Infection in Adults and Adolescents - UpToDateFer AguilarAún no hay calificaciones
- Diagnóstico Diferencial de Endocarditis InfecciosaDocumento3 páginasDiagnóstico Diferencial de Endocarditis InfecciosaFer AguilarAún no hay calificaciones
- Examen Economia en Salud 2020 Ii Ramos Aguilar Maria FernandaDocumento3 páginasExamen Economia en Salud 2020 Ii Ramos Aguilar Maria FernandaFer AguilarAún no hay calificaciones
- Endocarditis Infecciosa - Fundamentos de La Práctica, Antecedentes, FisiopatologíaDocumento18 páginasEndocarditis Infecciosa - Fundamentos de La Práctica, Antecedentes, FisiopatologíaFer AguilarAún no hay calificaciones
- Tratamiento y Manejo de La Endocarditis Infecciosa - Consideraciones de Abordaje, Terapia Con Antibióticos, Manejo de La Bacteriemia Por S. AureusDocumento14 páginasTratamiento y Manejo de La Endocarditis Infecciosa - Consideraciones de Abordaje, Terapia Con Antibióticos, Manejo de La Bacteriemia Por S. AureusFer AguilarAún no hay calificaciones
- Pautas para La Endocarditis Infecciosa - Resumen de Las PautasDocumento3 páginasPautas para La Endocarditis Infecciosa - Resumen de Las PautasFer AguilarAún no hay calificaciones
- Hepatitis - StatPearls - NCBI BookshelfDocumento31 páginasHepatitis - StatPearls - NCBI BookshelfFer AguilarAún no hay calificaciones
- Descripción General de Los Agentes Antirretrovirales Utilizados para Tratar El VIH - UpToDateDocumento45 páginasDescripción General de Los Agentes Antirretrovirales Utilizados para Tratar El VIH - UpToDateFer AguilarAún no hay calificaciones
- Infección Aguda y Temprana Por VIH - Manifestaciones Clínicas y Diagnóstico - UpToDateDocumento23 páginasInfección Aguda y Temprana Por VIH - Manifestaciones Clínicas y Diagnóstico - UpToDateFer AguilarAún no hay calificaciones
- Treatment of AIDS-related Cytomegalovirus Retinitis - UpToDateDocumento29 páginasTreatment of AIDS-related Cytomegalovirus Retinitis - UpToDateFer AguilarAún no hay calificaciones
- Cuándo Iniciar La Terapia Antirretroviral en Personas Con VIH - UpToDateDocumento14 páginasCuándo Iniciar La Terapia Antirretroviral en Personas Con VIH - UpToDateFer AguilarAún no hay calificaciones
- Patologías GenitourinarioDocumento1 páginaPatologías GenitourinarioFer AguilarAún no hay calificaciones
- Complejo Mycobacterium Avium - StatPearls - NCBI BookshelfDocumento9 páginasComplejo Mycobacterium Avium - StatPearls - NCBI BookshelfFer AguilarAún no hay calificaciones
- Hipertensión ArterialDocumento1 páginaHipertensión ArterialFer AguilarAún no hay calificaciones
- EPOCDocumento1 páginaEPOCFer AguilarAún no hay calificaciones
- Antidiabéticos OralesDocumento4 páginasAntidiabéticos OralesFer AguilarAún no hay calificaciones
- Diarrea AgudaDocumento1 páginaDiarrea AgudaFer AguilarAún no hay calificaciones
- Esquema de Reanimación NeonatalDocumento1 páginaEsquema de Reanimación NeonatalFer AguilarAún no hay calificaciones
- El Trabajo y Su Relacion Con La SaludDocumento13 páginasEl Trabajo y Su Relacion Con La SaludEliana Mariana RomeroAún no hay calificaciones
- Protocolo Ferroterapia IV v4Documento9 páginasProtocolo Ferroterapia IV v4IrianaAún no hay calificaciones
- Tesis Obtencion Del Titulo de OdontologoDocumento89 páginasTesis Obtencion Del Titulo de OdontologoChristopher Lopez ReyesAún no hay calificaciones
- Exposicion de Exploración Pediatrica eDocumento73 páginasExposicion de Exploración Pediatrica eConcepcion Sereno RosalesAún no hay calificaciones
- Disfunción Sacroilíaca y Dolor Lumbar Relacionado Con El Embarazo, Son Efectivas Las Terapias Médico-ManualesDocumento5 páginasDisfunción Sacroilíaca y Dolor Lumbar Relacionado Con El Embarazo, Son Efectivas Las Terapias Médico-ManualesDan CarcamoAún no hay calificaciones
- InfectologiíaDocumento74 páginasInfectologiíaMitchie AlbertiAún no hay calificaciones
- DSM TareaDocumento3 páginasDSM TareaHector VelizAún no hay calificaciones
- Sipnosis The SinnerDocumento4 páginasSipnosis The Sinnerenmanuel1209Aún no hay calificaciones
- Ultrasonografia ObstetricaDocumento7 páginasUltrasonografia ObstetricaLuchito CbaAún no hay calificaciones
- Rehabilitación de Un Paciente Desdentado TotalDocumento120 páginasRehabilitación de Un Paciente Desdentado Totalreyesking95Aún no hay calificaciones
- DigestionDocumento15 páginasDigestionNAYELI LORENA HUARZAYA CUPIAún no hay calificaciones
- Parasitología (IX) : Enfermedad de Chagas o Tripanosomiasis AmericanaDocumento89 páginasParasitología (IX) : Enfermedad de Chagas o Tripanosomiasis Americanaalfonso padillaAún no hay calificaciones
- Version Final Dediagram de Flujo RABIADocumento1 páginaVersion Final Dediagram de Flujo RABIABedlan BeckerAún no hay calificaciones
- Estrategias para El Diagnostico ClinicoDocumento15 páginasEstrategias para El Diagnostico Clinicodebbie38Aún no hay calificaciones
- Enfermedad Vascular CerebralDocumento50 páginasEnfermedad Vascular CerebralBlanca Aide LópezAún no hay calificaciones
- Las 8 Etapas Del Desarrollo HumanoDocumento2 páginasLas 8 Etapas Del Desarrollo Humanodarleny jimenezAún no hay calificaciones
- ALOTEXTO NapoleónDocumento2 páginasALOTEXTO NapoleónYury OlarteAún no hay calificaciones
- Fermentacion Liquida ShiitakeDocumento122 páginasFermentacion Liquida ShiitakeKatherineSerranoAún no hay calificaciones
- Dietaq Con YodoDocumento8 páginasDietaq Con YodoADDA HOYOSAún no hay calificaciones
- Asfixia Perinatal.Documento47 páginasAsfixia Perinatal.ingrydAún no hay calificaciones
- Temas Selectos en OtorrinolaringologiaDocumento377 páginasTemas Selectos en Otorrinolaringologiaangelica50% (2)
- Alteración Microbiana de Alientos Carnes Frescas Y Aves de CorralDocumento35 páginasAlteración Microbiana de Alientos Carnes Frescas Y Aves de CorralDavid Stiven RAún no hay calificaciones
- Bruno Latour - Dadme Un Laboratorio y Levantare El MundoDocumento20 páginasBruno Latour - Dadme Un Laboratorio y Levantare El MundoNinna Molano GranadosAún no hay calificaciones
- Células Sanguíneas...Documento1 páginaCélulas Sanguíneas...Andrea VenegasAún no hay calificaciones
- Lesiones Reactivas de La Cavidad BucalDocumento3 páginasLesiones Reactivas de La Cavidad BucalMartin GafeAún no hay calificaciones
- Suturas en OdontologiaDocumento28 páginasSuturas en OdontologiaVanesa Sigueñas Cajo100% (2)
- Plaguicidas Usados en BoliviaDocumento8 páginasPlaguicidas Usados en BoliviaHugo Uzeda AchúcarroAún no hay calificaciones