También podría gustarte
- Introducción a la Biología: RESÚMENES UNIVERSITARIOSDe EverandIntroducción a la Biología: RESÚMENES UNIVERSITARIOSCalificación: 5 de 5 estrellas5/5 (1)
- El Gran Camino Del Despertar - Jamgon KongtrulDocumento107 páginasEl Gran Camino Del Despertar - Jamgon KongtrulFran J. SANCHEZ100% (2)
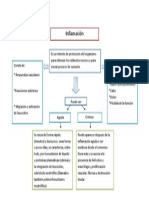
- Mapa Conceptual InflamacionDocumento1 páginaMapa Conceptual InflamacionveratosAún no hay calificaciones
- 1 5Documento4 páginas1 5gabyguillo50% (2)
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012De EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012Aún no hay calificaciones
- De Gen A Proteina-Inf11-SemDocumento7 páginasDe Gen A Proteina-Inf11-SemLuis Alejandro Quispe PonceAún no hay calificaciones
- FSSC 22000Documento65 páginasFSSC 22000Tacianus Velázquez80% (5)
- Grupos étnicos en ColombiaDocumento20 páginasGrupos étnicos en ColombiaOrlando Huertas100% (1)
- Retículo EndoplasmáticoDocumento36 páginasRetículo EndoplasmáticoMichael Morse100% (1)
- Informe S6.BIO - SEM-Grupo08Documento10 páginasInforme S6.BIO - SEM-Grupo08Kenner Poult Santisteban MuñozAún no hay calificaciones
- Cap11,12,13 Bio54Documento16 páginasCap11,12,13 Bio54Diego GaunaAún no hay calificaciones
- Nucleo 1Documento7 páginasNucleo 1Lau OrellanaAún no hay calificaciones
- Estructura Del Complejo Del Poro Nuclear-2Documento16 páginasEstructura Del Complejo Del Poro Nuclear-2Yinna Colina LugoAún no hay calificaciones
- TP 9 Membranas Internas IDocumento9 páginasTP 9 Membranas Internas ICaterina Maurin 8 6 3 7 5 / 4Aún no hay calificaciones
- Capitulo 15 Compartimientos y Transporte IntracelularDocumento6 páginasCapitulo 15 Compartimientos y Transporte IntracelularFlorangela MartinezAún no hay calificaciones
- Compartimentos Intracelulares y Clasificación de Proteínas PDFDocumento6 páginasCompartimentos Intracelulares y Clasificación de Proteínas PDFStanaAlehgnaAún no hay calificaciones
- Bloque Tematico 2 BiologiaDocumento39 páginasBloque Tematico 2 Biologiaangelica carrilloAún no hay calificaciones
- Compartimentación CelularDocumento3 páginasCompartimentación CelularFederico CalvoAún no hay calificaciones
- El Núcleo y CromosomaDocumento4 páginasEl Núcleo y CromosomaHeidy Villavicencio BoyerAún no hay calificaciones
- Expo SISTEMA MEMBRANASDocumento28 páginasExpo SISTEMA MEMBRANASVANIA ALEXA COTRINA ESTEVESAún no hay calificaciones
- BC 7 NecroticaenfmedDocumento8 páginasBC 7 NecroticaenfmedCamilaAún no hay calificaciones
- Biologia SEMANA 9Documento4 páginasBiologia SEMANA 9john salchichon67% (9)
- Ada 5 (BCM)Documento14 páginasAda 5 (BCM)Diana CetinaAún no hay calificaciones
- 11-Sistema de Endomembranas-Apunte GeneralDocumento19 páginas11-Sistema de Endomembranas-Apunte GeneralTomas Scialfa BenderskyAún no hay calificaciones
- 14-06-15 Genomasur Summary BiologíaDocumento6 páginas14-06-15 Genomasur Summary Biologíamodern_dudeAún no hay calificaciones
- Prueba n2 de BiologiaDocumento22 páginasPrueba n2 de BiologiaNohelis MelgarAún no hay calificaciones
- Clase 5-Biología - Gen - 24 - VeranoDocumento15 páginasClase 5-Biología - Gen - 24 - Veranoacostajohn397Aún no hay calificaciones
- Importación y Exportación en El Núcleo CelularDocumento4 páginasImportación y Exportación en El Núcleo CelularJardines del Rio EcuadorAún no hay calificaciones
- T.9 Compartimentos Intracelulares y Tránsito de PorteinasDocumento6 páginasT.9 Compartimentos Intracelulares y Tránsito de PorteinasNatalia Villar CrespoAún no hay calificaciones
- Clase 13 - Núcleo CelularDocumento9 páginasClase 13 - Núcleo CelularDulce SuárezAún no hay calificaciones
- NUCLEO CELULAR Cromatina y CromosomasDocumento44 páginasNUCLEO CELULAR Cromatina y CromosomasAbraham CutipaAún no hay calificaciones
- Retículo EndoplasmáticoDocumento20 páginasRetículo EndoplasmáticoAlba Macipe FeitoAún no hay calificaciones
- El NúcleoDocumento8 páginasEl NúcleoAlba Macipe FeitoAún no hay calificaciones
- Membranas InternasDocumento9 páginasMembranas InternasFranciscaCerdaAún no hay calificaciones
- Retículo EndoplasmáticoDocumento8 páginasRetículo Endoplasmáticocongaconga123Aún no hay calificaciones
- Proteínas sintetizadas: destinos y rutas celularesDocumento14 páginasProteínas sintetizadas: destinos y rutas celularesRita TolabaAún no hay calificaciones
- CLASIFICACIÓN PROTEÍNAS-DIRECCIONAMIENTODocumento31 páginasCLASIFICACIÓN PROTEÍNAS-DIRECCIONAMIENTOAshly MartínezAún no hay calificaciones
- Trafico CelularDocumento92 páginasTrafico Celularhalices0% (1)
- ESTRUCTURA Y FUNCIONES DEL NUCLEO CELULARDocumento36 páginasESTRUCTURA Y FUNCIONES DEL NUCLEO CELULARsabrina100% (1)
- Transporte IntracelularDocumento7 páginasTransporte IntracelularRamiro Santamaria100% (1)
- Compartimentos Intracelulares y Sorting de ProteínasDocumento9 páginasCompartimentos Intracelulares y Sorting de ProteínasFelipe Calderon AvendañoAún no hay calificaciones
- Unidad Didáctica IvDocumento10 páginasUnidad Didáctica Ivtrasteandopuntocom tpcAún no hay calificaciones
- Resumen 3er Modulo TeoriaDocumento10 páginasResumen 3er Modulo TeoriaCarrión Melo Alejandro MartinAún no hay calificaciones
- Biologia Celular Segundo ExamenDocumento11 páginasBiologia Celular Segundo ExamenCinthia BuchhammerAún no hay calificaciones
- EndosomasDocumento3 páginasEndosomasAriana ReyesAún no hay calificaciones
- Sistema de EndomembranasDocumento2 páginasSistema de EndomembranasFacu CorralesAún no hay calificaciones
- Biologia Celular Materia Solemne 3Documento5 páginasBiologia Celular Materia Solemne 3Alejandra MuñozAún no hay calificaciones
- Preguntas Cap 11Documento12 páginasPreguntas Cap 11Ariadni ChiquilloAún no hay calificaciones
- Guía Biologia CelularDocumento18 páginasGuía Biologia Celularalexa franciscaAún no hay calificaciones
- Exportación y Transporte de ProteinasDocumento27 páginasExportación y Transporte de Proteinasmiguel angel90% (10)
- Membranas Celulares InternasDocumento9 páginasMembranas Celulares InternasANA SIBAJAAún no hay calificaciones
- Biologia CelularDocumento114 páginasBiologia Celularbelem martinezAún no hay calificaciones
- 3 NucleoDocumento8 páginas3 NucleoDahiana BergesAún no hay calificaciones
- DSC5831Documento76 páginasDSC583120206394Aún no hay calificaciones
- Folio Nº8 Resumen Rayssa Xavier 2 - 221018 - 001618Documento40 páginasFolio Nº8 Resumen Rayssa Xavier 2 - 221018 - 001618LautaroAún no hay calificaciones
- Unidad 3 - Sistemas de EndomembranasDocumento17 páginasUnidad 3 - Sistemas de EndomembranasMelAún no hay calificaciones
- Copia de Resumen de HistologiaDocumento81 páginasCopia de Resumen de HistologiaAbigail DziubekAún no hay calificaciones
- Resumen Contenidos Unidad VIIDocumento7 páginasResumen Contenidos Unidad VIIALEJANDRO IGNACIO RUBIO ARENASAún no hay calificaciones
- El Núcleo Sirve de Almacén de La Información Genética y Como Centro de Control CelularDocumento2 páginasEl Núcleo Sirve de Almacén de La Información Genética y Como Centro de Control Celularangie c romeroAún no hay calificaciones
- Modulo III Reticulo Endoplásmicosistema de Endomembranas Con Audio Final 2023Documento29 páginasModulo III Reticulo Endoplásmicosistema de Endomembranas Con Audio Final 2023Madelin BolañosAún no hay calificaciones
- Resumen Segundo Parcial de BiologíaDocumento22 páginasResumen Segundo Parcial de BiologíaBelén barriosAún no hay calificaciones
- Estructura y función celularDocumento21 páginasEstructura y función celularagustin baccaroAún no hay calificaciones
- Sistema de EndomembranasDocumento27 páginasSistema de EndomembranasSofía TorresAún no hay calificaciones
- Membrana NuclearDocumento54 páginasMembrana NuclearLuis David Bello RodríguezAún no hay calificaciones
- Repaso Tercer Parcial Favaloro biologia 1ro medicinaDocumento3 páginasRepaso Tercer Parcial Favaloro biologia 1ro medicinaluizagustinboylerAún no hay calificaciones
- CITOESQUELETO.Documento12 páginasCITOESQUELETO.pimenteltamara2Aún no hay calificaciones
- Membrana celular: estructura y funcionesDocumento5 páginasMembrana celular: estructura y funcionesMazuey CroissantsAún no hay calificaciones
- Guia de estudio COMPARTIMIENTOS INTRACELULARESDocumento47 páginasGuia de estudio COMPARTIMIENTOS INTRACELULARESpimenteltamara2Aún no hay calificaciones
- SENALIZACION CELULARDocumento6 páginasSENALIZACION CELULARpimenteltamara2Aún no hay calificaciones
- Microscopia y TecnicasDocumento23 páginasMicroscopia y Tecnicaspimenteltamara2Aún no hay calificaciones
- RESÚMEN DE BIOFÍSICA 3Documento43 páginasRESÚMEN DE BIOFÍSICA 3pimenteltamara2Aún no hay calificaciones
- Coloides FQDocumento29 páginasColoides FQAbel YPAún no hay calificaciones
- Cotización Máquina Nueva Nacional Fitness StreamDocumento8 páginasCotización Máquina Nueva Nacional Fitness StreamEnmanuel Antony JeriAún no hay calificaciones
- Cámara frigorífica 50t espárragosDocumento4 páginasCámara frigorífica 50t espárragosCarlos Francisco LunaAún no hay calificaciones
- Aire AcondicionadoDocumento82 páginasAire AcondicionadoCarlos OchoaAún no hay calificaciones
- Cola Granulada PDFDocumento1 páginaCola Granulada PDFRoberto Toledo MolinaAún no hay calificaciones
- Práctica 1. Reconocimiento Del LaboratorioDocumento6 páginasPráctica 1. Reconocimiento Del LaboratorioGissela PumisachoAún no hay calificaciones
- EVALUACIÓN DE UNA TIERRA PARA EL APROVECHAMIENTO DE LA TRUFA NEGRA (Tuber Melanosporum Vittad.) EN ENCINAR: CASO TARIEGO DEL CERRATO, PALENCIA, ESPAÑADocumento53 páginasEVALUACIÓN DE UNA TIERRA PARA EL APROVECHAMIENTO DE LA TRUFA NEGRA (Tuber Melanosporum Vittad.) EN ENCINAR: CASO TARIEGO DEL CERRATO, PALENCIA, ESPAÑASamuel González BrantuasAún no hay calificaciones
- Actividad 1 Tema 1Documento5 páginasActividad 1 Tema 1Rolando RolesAún no hay calificaciones
- Areas de La Cocina PDFDocumento3 páginasAreas de La Cocina PDFPatricia Dorado44% (9)
- Guia Anatomia Humana (DBIO-1051)Documento268 páginasGuia Anatomia Humana (DBIO-1051)morenita1977Aún no hay calificaciones
- BoraxDocumento4 páginasBoraxDavid HoyosAún no hay calificaciones
- Quiz - Escenario 3 - SEGUNDO BLOQUE-TEORICO - VIRTUAL - CULTURA AMBIENTAL - (GRUPO B10)Documento8 páginasQuiz - Escenario 3 - SEGUNDO BLOQUE-TEORICO - VIRTUAL - CULTURA AMBIENTAL - (GRUPO B10)Maria fernanda G CubillosAún no hay calificaciones
- TALLER Tercer ParcialDocumento3 páginasTALLER Tercer ParcialSantiago RuedaAún no hay calificaciones
- Catalogo JuguetesDocumento94 páginasCatalogo JuguetesJosué AlviaAún no hay calificaciones
- Embriología - DESARROLLO DEL SISTEMA ÓSEO MUSCULAR DEL FETODocumento8 páginasEmbriología - DESARROLLO DEL SISTEMA ÓSEO MUSCULAR DEL FETOMiluska Rosario Pocco LozadaAún no hay calificaciones
- Taller Quimica OrganicaDocumento25 páginasTaller Quimica OrganicaSantiago España GutierrezAún no hay calificaciones
- El Amor y La InfidelidadDocumento27 páginasEl Amor y La InfidelidadJulissa Rosa Castro CastroAún no hay calificaciones
- Articulo Reovirus Aviar MaekawaDocumento9 páginasArticulo Reovirus Aviar MaekawaMariely Dennisse Segovia Alvarez0% (1)
- El MFC en BoliviaDocumento15 páginasEl MFC en BoliviajselAún no hay calificaciones
- Preguntas de FarmacologiaDocumento4 páginasPreguntas de FarmacologiadanielaAún no hay calificaciones
- Fisioterapeuta CV DuitamaDocumento15 páginasFisioterapeuta CV DuitamaJAVIER GONZALEZAún no hay calificaciones
- Copia de GUIA FINAL DE MICRODocumento19 páginasCopia de GUIA FINAL DE MICROXiomy LobattoAún no hay calificaciones
- Sesión 10Documento17 páginasSesión 10Abel LozanoAún no hay calificaciones
- Contabilidad de Costos 1Documento3 páginasContabilidad de Costos 1ElizabethAún no hay calificaciones
- Servicio PediatriaDocumento8 páginasServicio PediatriaMaria Luisa DelgadoAún no hay calificaciones