También podría gustarte
- Introducción a la Biología: RESÚMENES UNIVERSITARIOSDe EverandIntroducción a la Biología: RESÚMENES UNIVERSITARIOSCalificación: 5 de 5 estrellas5/5 (1)
- Crecimiento BacterianoDocumento58 páginasCrecimiento BacterianoNitgma DcAún no hay calificaciones
- Ciclo Celular ProcarióticoDocumento11 páginasCiclo Celular Procarióticoatomo5250% (6)
- Fases Del Ciclo CelularDocumento5 páginasFases Del Ciclo CelularMica VargasAún no hay calificaciones
- Ciclo Celular ProcariotaDocumento25 páginasCiclo Celular ProcariotaPriscilla Mulla Lazo0% (1)
- Taller 5 de Génetica CáncerDocumento16 páginasTaller 5 de Génetica CáncerMaría MontoyaAún no hay calificaciones
- Trabajo Ciclo CelularDocumento16 páginasTrabajo Ciclo CelularBeatriz Cervantes AlvaradoAún no hay calificaciones
- Tema 3 División CelularDocumento23 páginasTema 3 División Celularjose juan mendezAún no hay calificaciones
- El Ciclo Celular y La ApoptosisDocumento19 páginasEl Ciclo Celular y La ApoptosisAlan OjedaAún no hay calificaciones
- Compendio Embrio Ciclo Celular y GametogenesisDocumento11 páginasCompendio Embrio Ciclo Celular y GametogenesisAstrid SofíaAún no hay calificaciones
- C8 División Ciclo Celular, Mitosis y MeiosisDocumento38 páginasC8 División Ciclo Celular, Mitosis y MeiosisNelida melendrezAún no hay calificaciones
- Ciclo CelularDocumento32 páginasCiclo CelularAnel Terranova100% (2)
- Taller Ciclo Celular NuevoDocumento6 páginasTaller Ciclo Celular NuevoDaniela Galvis HernandezAún no hay calificaciones
- Separata Ciclo CelularDocumento11 páginasSeparata Ciclo CelularJohn Antony Vilchez RetamozoAún no hay calificaciones
- Seminario de CancerDocumento28 páginasSeminario de CancerKevin García Murga100% (1)
- Ciclo CelularDocumento3 páginasCiclo CelularMaría Celeste PascualAún no hay calificaciones
- Apuntes - El Ciclo CelularDocumento17 páginasApuntes - El Ciclo CelularGaby MateosAún no hay calificaciones
- Ciclo Celular Guia 1Documento9 páginasCiclo Celular Guia 1Belen MercadoAún no hay calificaciones
- Machete 7Documento27 páginasMachete 744315038Aún no hay calificaciones
- Guia Pràctica Laboratorio Sesión 11Documento8 páginasGuia Pràctica Laboratorio Sesión 11Kevin JoseAún no hay calificaciones
- Taller 28Documento7 páginasTaller 28santiago morenoAún no hay calificaciones
- Guia Aprendizaje Division CelularDocumento11 páginasGuia Aprendizaje Division CelularJuan Manuel Neira Ferrada0% (1)
- Qué Es El Ciclo CelularDocumento5 páginasQué Es El Ciclo CelularSu SaucedoAún no hay calificaciones
- Ciclo CelularDocumento6 páginasCiclo CelularMicaela CostillaAún no hay calificaciones
- Paper de Ciclo Celular.Documento16 páginasPaper de Ciclo Celular.Jean Carlos Hernandez BeltranAún no hay calificaciones
- Taller Ciclo Celular B. GeneralDocumento4 páginasTaller Ciclo Celular B. GeneralDavid Vergara MorenoAún no hay calificaciones
- Resumen 2Documento4 páginasResumen 2xochitl palma naranjoAún no hay calificaciones
- Ciclo Celular y Duplicación Del AdnDocumento60 páginasCiclo Celular y Duplicación Del AdnDonald MartinAún no hay calificaciones
- Niversidad Privada Antenor Orrego BiologDocumento28 páginasNiversidad Privada Antenor Orrego BiologEsmeralda LopezAún no hay calificaciones
- Ciclo CelularDocumento3 páginasCiclo CelularAngel Valdez ArreolaAún no hay calificaciones
- Ciclo CelularDocumento8 páginasCiclo Celularana villotaAún no hay calificaciones
- Guia Ciclo Celular (Mitosis) Meiosis y Gametogenesis EnfermeriaDocumento27 páginasGuia Ciclo Celular (Mitosis) Meiosis y Gametogenesis EnfermeriaGhillian Amanda Doll AlucemaAún no hay calificaciones
- Wa0023. - 083520Documento12 páginasWa0023. - 083520Rafael Comesaña LealAún no hay calificaciones
- Ciclo CelularDocumento23 páginasCiclo CelularOrtiz Alba AndreaAún no hay calificaciones
- Unidad 7. Mitosis y Meiosis 2023Documento62 páginasUnidad 7. Mitosis y Meiosis 2023Alan GanemAún no hay calificaciones
- Lectura 5-Division Celular y MeiosisDocumento5 páginasLectura 5-Division Celular y Meiosislizzy510Aún no hay calificaciones
- Taller Genetica ADNDocumento9 páginasTaller Genetica ADNTefi OspinaAún no hay calificaciones
- Ciclo CelularDocumento5 páginasCiclo CelularJuan MendozaAún no hay calificaciones
- Ciclo CelularDocumento11 páginasCiclo CelularEdward Balvin SincheAún no hay calificaciones
- Division CelularDocumento11 páginasDivision CelularDiana ReinalesAún no hay calificaciones
- U05 M01 Ciclo CelularDocumento5 páginasU05 M01 Ciclo CelularSanlucasAún no hay calificaciones
- Semana 12 Ciclo Celular y MitosisDocumento18 páginasSemana 12 Ciclo Celular y MitosisShantal Ponce JáureguiAún no hay calificaciones
- Ciclo CelularDocumento11 páginasCiclo Celulardavid eduardo lopez quirozAún no hay calificaciones
- Monografia de Biologia Cilco CelularDocumento12 páginasMonografia de Biologia Cilco Celularibeth rincon duranAún no hay calificaciones
- Seminario 2 Ciclo Celular y Cancer 2023-IDocumento28 páginasSeminario 2 Ciclo Celular y Cancer 2023-IMariafe PurizagaAún no hay calificaciones
- Apuntes Ciclo CelularDocumento24 páginasApuntes Ciclo CelularGaby CaroAún no hay calificaciones
- INFORME S9 Biologia PracticaDocumento13 páginasINFORME S9 Biologia PracticaMarycielo MmsAún no hay calificaciones
- GUIA DE SEMINARIO CICLO CELULAR FinalDocumento11 páginasGUIA DE SEMINARIO CICLO CELULAR FinalUrania MoyaAún no hay calificaciones
- Bloque VDocumento10 páginasBloque VEffectdZebraAún no hay calificaciones
- Ciclo CelularDocumento9 páginasCiclo CelularKent Junior Limas AyalaAún no hay calificaciones
- Ciclo Celular EucariotaDocumento3 páginasCiclo Celular Eucariotapaola laraAún no hay calificaciones
- Ficha de Estudio Reproduccion CelularDocumento3 páginasFicha de Estudio Reproduccion CelularAmanecer ImpresosAún no hay calificaciones
- Clase 1. GENETICA (MARIA GABRIELA RIVAS)Documento14 páginasClase 1. GENETICA (MARIA GABRIELA RIVAS)lobogonzalezjAún no hay calificaciones
- División Celular, Mitosis y MeiosisDocumento8 páginasDivisión Celular, Mitosis y MeiosisMariana ReyesAún no hay calificaciones
- Ciclo Celular - KHP - TC01Documento12 páginasCiclo Celular - KHP - TC01Kenia HernandezAún no hay calificaciones
- 6 Biologiageneticaysociedad Actividad5Documento7 páginas6 Biologiageneticaysociedad Actividad5Vale MonteroAún no hay calificaciones
- LADocumento51 páginasLATomoyo SakuraAún no hay calificaciones
- Documento PDFDocumento1 páginaDocumento PDFmargarita oshea gumucioAún no hay calificaciones
- Ciclo Celular Puntos de Control Del Ciclo Celular Flujo de Información Genética Ácidos NucleicosDocumento29 páginasCiclo Celular Puntos de Control Del Ciclo Celular Flujo de Información Genética Ácidos NucleicosGonzalo LeoAún no hay calificaciones
- Unida-2 Semana 10 Clase 1 LunesDocumento65 páginasUnida-2 Semana 10 Clase 1 Lunesbriyith garciaAún no hay calificaciones
- DoclazekmduDocumento5 páginasDoclazekmduVrex KingAún no hay calificaciones
- Genética ExposiciónDocumento3 páginasGenética ExposiciónVrex KingAún no hay calificaciones
- Protozoo4 DefDocumento47 páginasProtozoo4 DefVrex KingAún no hay calificaciones
- 6 Diferencias Del ADN y ARNDocumento1 página6 Diferencias Del ADN y ARNVrex KingAún no hay calificaciones
- CholinesteraseDocumento16 páginasCholinesteraseVrex KingAún no hay calificaciones
- AnimalesDocumento7 páginasAnimalesVrex KingAún no hay calificaciones
- Trabajo paraDocumento20 páginasTrabajo paraVrex KingAún no hay calificaciones
- DiosDocumento14 páginasDiosVrex KingAún no hay calificaciones
- 6 Diferencias Del ADN y ARNDocumento1 página6 Diferencias Del ADN y ARNVrex KingAún no hay calificaciones
- Evaluacion Unidad 3Documento2 páginasEvaluacion Unidad 3Vrex KingAún no hay calificaciones
- Tipos de Respiración: © 2013-2023 Enciclopedia Concepto. Todos Los Derechos ReservadosDocumento12 páginasTipos de Respiración: © 2013-2023 Enciclopedia Concepto. Todos Los Derechos ReservadosVrex KingAún no hay calificaciones
- Retículo Sarcoplásmico: Publicado 25 de Mayo de 2015, 14:39Documento5 páginasRetículo Sarcoplásmico: Publicado 25 de Mayo de 2015, 14:39Vrex KingAún no hay calificaciones
- 2 Actividad Genetica 2024 Grupo 5Documento1 página2 Actividad Genetica 2024 Grupo 5Vrex KingAún no hay calificaciones
- Guia de Metabolismo D LipidosDocumento9 páginasGuia de Metabolismo D LipidosVenezuelaAún no hay calificaciones
- MalinalcoDocumento4 páginasMalinalcololibelcgAún no hay calificaciones
- Retículo Sarcoplásmico: Publicado 25 de Mayo de 2015, 14:39Documento5 páginasRetículo Sarcoplásmico: Publicado 25 de Mayo de 2015, 14:39Vrex KingAún no hay calificaciones
- DkjejejsoDocumento5 páginasDkjejejsoVrex KingAún no hay calificaciones
- Procesos y Tipos de RespiraciónDocumento5 páginasProcesos y Tipos de RespiraciónVrex KingAún no hay calificaciones
- Documento (1) BBVDHDVDocumento6 páginasDocumento (1) BBVDHDVVrex KingAún no hay calificaciones
- DkjejejsoDocumento5 páginasDkjejejsoVrex KingAún no hay calificaciones
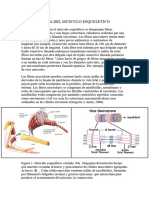
- Ii. Morfologia Del Musculo EsqueleticoDocumento21 páginasIi. Morfologia Del Musculo EsqueleticoVrex KingAún no hay calificaciones
- Tipos de Respiración: © 2013-2023 Enciclopedia Concepto. Todos Los Derechos ReservadosDocumento12 páginasTipos de Respiración: © 2013-2023 Enciclopedia Concepto. Todos Los Derechos ReservadosVrex KingAún no hay calificaciones
- 98.6 F98.6 F 37 C37 C: Puntos Más ImportantesDocumento9 páginas98.6 F98.6 F 37 C37 C: Puntos Más ImportantesVrex KingAún no hay calificaciones
- Tema3 Protozoo5Documento22 páginasTema3 Protozoo5Vrex KingAún no hay calificaciones
- Guía Practica de Parasitología Sistematica Zoologica y Protozoos1Documento34 páginasGuía Practica de Parasitología Sistematica Zoologica y Protozoos1Vrex KingAún no hay calificaciones
- Guía Practica de Parasitología Sistematica Zoologica y Protozoos1Documento34 páginasGuía Practica de Parasitología Sistematica Zoologica y Protozoos1Vrex KingAún no hay calificaciones
- Bacterias 2Documento6 páginasBacterias 2paula noemi argañarazAún no hay calificaciones
- Ii. Morfologia Del Musculo EsqueleticoDocumento21 páginasIi. Morfologia Del Musculo EsqueleticoVrex KingAún no hay calificaciones
- Biomoléculas Orgánicas o Principios InmediatosDocumento4 páginasBiomoléculas Orgánicas o Principios InmediatosVrex KingAún no hay calificaciones
- Tema1 2Documento23 páginasTema1 2Vrex KingAún no hay calificaciones
- 2019 10 10 Espinosa Barrera y Chavez Sahagun El Otro Genoma de Las Plantas PDFDocumento6 páginas2019 10 10 Espinosa Barrera y Chavez Sahagun El Otro Genoma de Las Plantas PDFAndrés Felipe Fernández QuirósAún no hay calificaciones
- VIRBAC - INMUNOMODULADORES BovinoslecheDocumento8 páginasVIRBAC - INMUNOMODULADORES BovinoslecheRobinho RicardzAún no hay calificaciones
- Prueba 1° Medio BiologíaDocumento5 páginasPrueba 1° Medio BiologíaVeronica NegreteAún no hay calificaciones
- Tansporte Pasivo y Transporte ActivoDocumento8 páginasTansporte Pasivo y Transporte ActivoAnonymous 4T2Qbd4Aún no hay calificaciones
- Membrana PlasmaticaDocumento5 páginasMembrana PlasmaticaCarolina PepeAún no hay calificaciones
- Sustratos SuicidasDocumento64 páginasSustratos SuicidasRuben Borda FloresAún no hay calificaciones
- Principios de Bioenergética 1a ClaseDocumento33 páginasPrincipios de Bioenergética 1a ClaseGabriel GuerreroAún no hay calificaciones
- Archea Bacteria (Caracteristicas Generales)Documento4 páginasArchea Bacteria (Caracteristicas Generales)Martin FigueroaAún no hay calificaciones
- Activacion de EnzimascDocumento3 páginasActivacion de EnzimascIsabel GasparAún no hay calificaciones
- RESUMEN Neuro COMPLETO!Documento160 páginasRESUMEN Neuro COMPLETO!aldanaAún no hay calificaciones
- Membrana CelularDocumento11 páginasMembrana CelulardianisssuxAún no hay calificaciones
- Radicales LibresDocumento3 páginasRadicales LibresRodrigo VillanuevaAún no hay calificaciones
- Expo Ciclo CelularDocumento27 páginasExpo Ciclo CelularNancyChrisJimenezAún no hay calificaciones
- Ensayo Sobre Los Niveles de Organización de La VidaDocumento3 páginasEnsayo Sobre Los Niveles de Organización de La VidaMirianny MeloAún no hay calificaciones
- Examen Olimpiada 2012Documento20 páginasExamen Olimpiada 2012Angela Lobato MañanesAún no hay calificaciones
- Respuestas FGen 2016-17Documento13 páginasRespuestas FGen 2016-17Romina SalazarAún no hay calificaciones
- Histo ClaseDocumento14 páginasHisto ClaseAdrian MendozaAún no hay calificaciones
- BIOLOGÍA Plan 1994 Examen Modelo 4Documento2 páginasBIOLOGÍA Plan 1994 Examen Modelo 4psico1322Aún no hay calificaciones
- Descarga PDFDocumento9 páginasDescarga PDFIvàn Luna GutierrezAún no hay calificaciones
- Sílabo Embriología Área 7 2017 - 2sDocumento16 páginasSílabo Embriología Área 7 2017 - 2sMiguel SagalAún no hay calificaciones
- 9examen Final Bases Biologicas y Antropologicas de La VidaDocumento15 páginas9examen Final Bases Biologicas y Antropologicas de La VidaCamila DiniAún no hay calificaciones
- Practica de CelulaDocumento8 páginasPractica de CelulaCamilo Andres Barreiro HerreraAún no hay calificaciones
- Topoisomerasa IIDocumento4 páginasTopoisomerasa IIJulian RiosAún no hay calificaciones
- Bio. Ácidos Nucleicos IiDocumento35 páginasBio. Ácidos Nucleicos IiJUANA ROSALIA GALVEZ MONCADAAún no hay calificaciones
- Membrana Plasmática Grupo 4Documento1 páginaMembrana Plasmática Grupo 4Ximena Ramirez100% (2)
- Ciclo Celular PDFDocumento8 páginasCiclo Celular PDFPablo Jorquera GarcésAún no hay calificaciones
- Laboratorio de HematologíaDocumento47 páginasLaboratorio de HematologíaMaria Isabel Ortiz TorresAún no hay calificaciones
- Sistema Vacuolar Citoplasmático1Documento52 páginasSistema Vacuolar Citoplasmático1Rodrigo RodriguezAún no hay calificaciones
- Lipoproteinas Analisis p.3Documento12 páginasLipoproteinas Analisis p.3jkr210% (1)
- Biologia InformeDocumento8 páginasBiologia Informecarlos9santacruz-2Aún no hay calificaciones
- GuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisDe EverandGuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisCalificación: 4 de 5 estrellas4/5 (9)
- Notas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesDe EverandNotas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesAún no hay calificaciones
- Nuestra mente nos engaña: Sesgos y errores cognitivos que todos cometemosDe EverandNuestra mente nos engaña: Sesgos y errores cognitivos que todos cometemosCalificación: 3.5 de 5 estrellas3.5/5 (6)
- 200 tareas en terapia breve: 2ª ediciónDe Everand200 tareas en terapia breve: 2ª ediciónCalificación: 4.5 de 5 estrellas4.5/5 (33)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- Abrázame fuerte: Siete conversaciones para un amor duraderoDe EverandAbrázame fuerte: Siete conversaciones para un amor duraderoManu BerásteguiCalificación: 4.5 de 5 estrellas4.5/5 (13)
- Neuropsicología: Los fundamentos de la materiaDe EverandNeuropsicología: Los fundamentos de la materiaCalificación: 5 de 5 estrellas5/5 (1)
- El cerebro del niño explicado a los padresDe EverandEl cerebro del niño explicado a los padresCalificación: 4.5 de 5 estrellas4.5/5 (147)
- Minerales y vitaminas: Micronutrientes esenciales en la alimentación, nutrición y salud.De EverandMinerales y vitaminas: Micronutrientes esenciales en la alimentación, nutrición y salud.Calificación: 3 de 5 estrellas3/5 (2)
- Cerebros rotos: Pacientes asombrosos que me enseñaron a vivirDe EverandCerebros rotos: Pacientes asombrosos que me enseñaron a vivirCalificación: 5 de 5 estrellas5/5 (3)
- El autismo: Reflexiones y pautas para comprenderlo y abordarloDe EverandEl autismo: Reflexiones y pautas para comprenderlo y abordarloCalificación: 4 de 5 estrellas4/5 (7)
- El perfil psicológico de Jesús: Aprendamos del Maestro a manejar efectivamente nuestras emocionesDe EverandEl perfil psicológico de Jesús: Aprendamos del Maestro a manejar efectivamente nuestras emocionesCalificación: 4 de 5 estrellas4/5 (4)
- La increíble conexión intestino cerebro: Descubre la relación entre las emociones y el equilibrio intestinalDe EverandLa increíble conexión intestino cerebro: Descubre la relación entre las emociones y el equilibrio intestinalCalificación: 4.5 de 5 estrellas4.5/5 (102)
- Cómo dormir bien: Cómo reconocer, prevenir y tratar las dificultades para dormir bienDe EverandCómo dormir bien: Cómo reconocer, prevenir y tratar las dificultades para dormir bienCalificación: 5 de 5 estrellas5/5 (1)
- Los cinco ritos tibetanos de la eterna juventud: El ojo de la revelaciónDe EverandLos cinco ritos tibetanos de la eterna juventud: El ojo de la revelaciónCalificación: 4.5 de 5 estrellas4.5/5 (12)
- Sesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesDe EverandSesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesCalificación: 4.5 de 5 estrellas4.5/5 (13)
- Resumen de Pensar rápido pensar despacio de Daniel KahnemanDe EverandResumen de Pensar rápido pensar despacio de Daniel KahnemanCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Principios básicos de bioquímica de los alimentosDe EverandPrincipios básicos de bioquímica de los alimentosCalificación: 4.5 de 5 estrellas4.5/5 (2)
- Fundamentos de procesos químicosDe EverandFundamentos de procesos químicosCalificación: 5 de 5 estrellas5/5 (3)
- Lentes de contacto: composición química y propiedadesDe EverandLentes de contacto: composición química y propiedadesCalificación: 3 de 5 estrellas3/5 (2)