También podría gustarte
- Gabriel Hernandez Fuentes Respuesta InmunitariaDocumento6 páginasGabriel Hernandez Fuentes Respuesta InmunitariaGabriel HernándezAún no hay calificaciones
- Taller Sars Cov 2 InmunoDocumento11 páginasTaller Sars Cov 2 InmunoEri MñzAún no hay calificaciones
- Receptores Tipo Toll - TareasDocumento12 páginasReceptores Tipo Toll - TareasDayana Torrico VAún no hay calificaciones
- Receptores Tipo Toll - TareasDocumento12 páginasReceptores Tipo Toll - TareasDayana Torrico VAún no hay calificaciones
- Inmunopatología - Trastorno de La HipersensibilidadDocumento5 páginasInmunopatología - Trastorno de La HipersensibilidadVanessa AvilaAún no hay calificaciones
- Inmunologia de LepraDocumento8 páginasInmunologia de LepraEfrain Teran CarbonoAún no hay calificaciones
- Conclusion EsDocumento2 páginasConclusion EsTito CollaguazoAún no hay calificaciones
- BRUCELLADocumento5 páginasBRUCELLAJhenny Orosco SolaresAún no hay calificaciones
- Estallido Respiratorio TareaDocumento5 páginasEstallido Respiratorio TareaAlain BallinasAún no hay calificaciones
- Inmunidad A HongosDocumento57 páginasInmunidad A HongosAndrés MesíaAún no hay calificaciones
- Inmunology TuberculosisDocumento50 páginasInmunology Tuberculosisfrancisco salazar huamaniAún no hay calificaciones
- TuberculosisDocumento5 páginasTuberculosisJane CertifyAún no hay calificaciones
- Inmunopatogenia de La TuberculosisDocumento14 páginasInmunopatogenia de La TuberculosisBrayan CcAún no hay calificaciones
- Inmunidad Contra Microorganismos - LópezlópezDocumento7 páginasInmunidad Contra Microorganismos - Lópezlópezdana lopezAún no hay calificaciones
- Cuestionario Respuesta InmunitariaDocumento5 páginasCuestionario Respuesta InmunitariaMercedes CamAún no hay calificaciones
- Fisiopatología de La SarcoidosisDocumento12 páginasFisiopatología de La SarcoidosisClaudioMolinaAún no hay calificaciones
- Inmunidad Frente A HongosDocumento19 páginasInmunidad Frente A Hongosjorge_chavez_189Aún no hay calificaciones
- Respuesta Inmunológica Frente A HongosDocumento2 páginasRespuesta Inmunológica Frente A HongosAlondra VegaAún no hay calificaciones
- Mecanismos Efectores de La Inmunidad AdaptativaDocumento8 páginasMecanismos Efectores de La Inmunidad AdaptativaEmilis Madrid100% (1)
- Patogenia HistoplasmosisDocumento6 páginasPatogenia HistoplasmosisCarlos Eduardo Celis SeclénAún no hay calificaciones
- 7 Activación-Lb-Y-LtDocumento14 páginas7 Activación-Lb-Y-LtGuille OliveraAún no hay calificaciones
- Patogenia de C.trachomatis y H.ducreyiDocumento2 páginasPatogenia de C.trachomatis y H.ducreyiarmando perez hernandezAún no hay calificaciones
- SRIS y Sepsis Artículo ResumenDocumento8 páginasSRIS y Sepsis Artículo ResumenMercedes CamAún no hay calificaciones
- Información de Toda La Exposición de InmunoDocumento5 páginasInformación de Toda La Exposición de InmunoJorge Andres Yanes ContrerasAún no hay calificaciones
- Clase INMUNIDAD FRENTE A VIRUS pt3Documento5 páginasClase INMUNIDAD FRENTE A VIRUS pt3Lucy Milagros LeónAún no hay calificaciones
- Ehrlichia CanisDocumento5 páginasEhrlichia CanisNaruto ShikamaruAún no hay calificaciones
- Inm-MOD 2Documento10 páginasInm-MOD 2NicoleAún no hay calificaciones
- Wuolah Free Tema 11 Gulag FreeDocumento10 páginasWuolah Free Tema 11 Gulag FreeY UuAún no hay calificaciones
- Produccion de Efectores Clase IIIDocumento71 páginasProduccion de Efectores Clase IIIOlga Meléndez MayorquinAún no hay calificaciones
- Efectores de La Inmunodad AdquiridaDocumento43 páginasEfectores de La Inmunodad AdquiridaOmar PopocaAún no hay calificaciones
- Casos de InmunologiaDocumento12 páginasCasos de InmunologiaVane Karen Rad AmudioAún no hay calificaciones
- La Inmunidad Innata A Toxoplasma Gondii InfecciónDocumento15 páginasLa Inmunidad Innata A Toxoplasma Gondii InfecciónRosario Sanchez SarmientoAún no hay calificaciones
- Respuesta Inflamatoria y AntiviralDocumento2 páginasRespuesta Inflamatoria y AntiviralIvan PachecoAún no hay calificaciones
- Agente Etiológico VihDocumento4 páginasAgente Etiológico VihKaren VeraAún no hay calificaciones
- Tipo de Comunicación Celular en La SepsisDocumento3 páginasTipo de Comunicación Celular en La SepsisluisamariaAún no hay calificaciones
- Grupo 20 - Inmunologia Seminario Informe S7Documento11 páginasGrupo 20 - Inmunologia Seminario Informe S7Dante ReevesAún no hay calificaciones
- IntroducciónDocumento20 páginasIntroducciónMauri GuañunaAún no hay calificaciones
- Inmunología de TuberculosisDocumento5 páginasInmunología de TuberculosisJessica ZendejasAún no hay calificaciones
- Citoquinas As Por Linfocitos Cd4Documento9 páginasCitoquinas As Por Linfocitos Cd4Claudia RodriguezAún no hay calificaciones
- Nos Ubicamos Segundo ParcialDocumento17 páginasNos Ubicamos Segundo ParcialBruno Masci MiralAún no hay calificaciones
- 169 Sistema Inmunitario Cutáneo PDFDocumento63 páginas169 Sistema Inmunitario Cutáneo PDFJ C Torres Formalab100% (1)
- Fisiopatologia Del DengueDocumento2 páginasFisiopatologia Del DenguePaola Andrea Diaz BermeoAún no hay calificaciones
- Inmunidad Frente A Bacterias Intracelulares - SalmonellaDocumento9 páginasInmunidad Frente A Bacterias Intracelulares - SalmonellaBrian Misari AnchiraicoAún no hay calificaciones
- Respuesta Inmunitaria Frente A VirusDocumento11 páginasRespuesta Inmunitaria Frente A VirusMaria Angeles Medina LlerenaAún no hay calificaciones
- El Papel de Los TLR en La Activación de NeutrófilosDocumento6 páginasEl Papel de Los TLR en La Activación de NeutrófilosVic VargasAún no hay calificaciones
- Candida Albicans Es Una Levadura Comensal Que Reside en Las Membranas Mucosas de Las Cavidades Oral y VaginalDocumento4 páginasCandida Albicans Es Una Levadura Comensal Que Reside en Las Membranas Mucosas de Las Cavidades Oral y VaginalDaniel CastilloAún no hay calificaciones
- Candida Albicans Es Una Levadura Comensal Que Reside en Las Membranas Mucosas de Las Cavidades Oral y VaginalDocumento4 páginasCandida Albicans Es Una Levadura Comensal Que Reside en Las Membranas Mucosas de Las Cavidades Oral y VaginalDaniel CastilloAún no hay calificaciones
- 1P Guía 4 Rpta InmuneDocumento3 páginas1P Guía 4 Rpta InmuneKaren AlloAún no hay calificaciones
- Captura de Pantalla 2022-03-22 A La(s) 11.46.23 A. M.Documento90 páginasCaptura de Pantalla 2022-03-22 A La(s) 11.46.23 A. M.Naila Di Trani HaitterAún no hay calificaciones
- Inmunologia 1Documento33 páginasInmunologia 1Kiara Antonia Quispe RamosAún no hay calificaciones
- Tuberculosis ResumenDocumento5 páginasTuberculosis ResumenOmar AdalidAún no hay calificaciones
- Clase INMUNIDAD FRENTE A VIRUS pt1Documento3 páginasClase INMUNIDAD FRENTE A VIRUS pt1Lucy Milagros LeónAún no hay calificaciones
- Celulas THDocumento5 páginasCelulas THmonse_moonAún no hay calificaciones
- Inmunologia Del DengueDocumento4 páginasInmunologia Del DengueAlessandra RomeroAún no hay calificaciones
- A Resposta Inmunitaria Frente A ParásitosDocumento5 páginasA Resposta Inmunitaria Frente A Parásitoscolordeestrellas92Aún no hay calificaciones
- PATOGENIADocumento2 páginasPATOGENIARaquel Noemi Pizan YupanquiAún no hay calificaciones
- 1C 03-02 Linfopoyesis - Monopoyesis - EritropoyesisDocumento55 páginas1C 03-02 Linfopoyesis - Monopoyesis - EritropoyesisEdward GilAún no hay calificaciones
- Función de Los MacrófagosDocumento3 páginasFunción de Los MacrófagosdanielaAún no hay calificaciones
- CEREBELITISDocumento27 páginasCEREBELITISKassandraAltamirano100% (1)
- Oficio Exoneración de ResponsabilidadDocumento1 páginaOficio Exoneración de ResponsabilidadKassandraAltamiranoAún no hay calificaciones
- Prof. Jorge Damián Foronda USP Chimbote-2014Documento60 páginasProf. Jorge Damián Foronda USP Chimbote-2014KassandraAltamiranoAún no hay calificaciones
- DIVERTICULOSISDocumento16 páginasDIVERTICULOSISKassandraAltamiranoAún no hay calificaciones
- Quintanilla Albán, Mayra BeatrizDocumento101 páginasQuintanilla Albán, Mayra BeatrizKassandraAltamiranoAún no hay calificaciones
- Endometriosis MonografiaDocumento36 páginasEndometriosis MonografiaKassandraAltamiranoAún no hay calificaciones
- Definition Epidemiology and Etiology of Obesity in Children and Adolescents UpToDateDocumento46 páginasDefinition Epidemiology and Etiology of Obesity in Children and Adolescents UpToDateKassandraAltamiranoAún no hay calificaciones
- Vol Vu Los Gastrointestinal EsDocumento17 páginasVol Vu Los Gastrointestinal EsKassandraAltamiranoAún no hay calificaciones
- Aspecto ReligiosoDocumento1 páginaAspecto ReligiosoKassandraAltamiranoAún no hay calificaciones
- BRONQUIECTASIASDocumento16 páginasBRONQUIECTASIASKassandraAltamirano50% (2)
- Rm0131-85-Sa-Adicion de Fluor A SalDocumento3 páginasRm0131-85-Sa-Adicion de Fluor A SalKassandraAltamiranoAún no hay calificaciones
- PARKINSONDocumento12 páginasPARKINSONKassandraAltamirano100% (2)
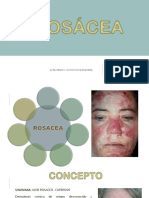
- ROSÁCEADocumento9 páginasROSÁCEAKassandraAltamiranoAún no hay calificaciones
- Bioestadistica Clase 2 Presentación1Documento79 páginasBioestadistica Clase 2 Presentación1KassandraAltamiranoAún no hay calificaciones
- Insuficiencia SuprarrenalDocumento7 páginasInsuficiencia SuprarrenalKassandraAltamiranoAún no hay calificaciones
- Bioestadistica y Demografia Clase 1 13 03 2015 Poblacion y MuestraDocumento48 páginasBioestadistica y Demografia Clase 1 13 03 2015 Poblacion y MuestraKassandraAltamiranoAún no hay calificaciones
- Potencia y Dosis Equivalentes de Los Glucocorticoides1 PDFDocumento1 páginaPotencia y Dosis Equivalentes de Los Glucocorticoides1 PDFKassandraAltamiranoAún no hay calificaciones
- Dictamen de Criminalistica2Documento4 páginasDictamen de Criminalistica2Vago Canto ContrerasAún no hay calificaciones
- Presentacion Acerca de La Pulpa Dental-Grupo BDocumento14 páginasPresentacion Acerca de La Pulpa Dental-Grupo BTania JudithAún no hay calificaciones
- Tema 52 - Actividad Física y SaludDocumento8 páginasTema 52 - Actividad Física y SaludJose Luis Millet Sanchez de la FuenteAún no hay calificaciones
- PumaDocumento4 páginasPumaKike InoztrozaAún no hay calificaciones
- Metamidofos FichaDocumento2 páginasMetamidofos FichaSalvadorMartinezToscanoAún no hay calificaciones
- Absorción de Fármacos PDFDocumento45 páginasAbsorción de Fármacos PDFJulieta MoraAún no hay calificaciones
- GENETICADocumento7 páginasGENETICAAlejandra ArriolaAún no hay calificaciones
- Relajación Progresiva de JacobsonDocumento12 páginasRelajación Progresiva de JacobsonSara GRAún no hay calificaciones
- KasapataDocumento6 páginasKasapataguillermo ttito sullcapumaAún no hay calificaciones
- CESTODOSDocumento56 páginasCESTODOSYINETH SOFIA VASQUEZ ROMEROAún no hay calificaciones
- Prueba de CienciasDocumento4 páginasPrueba de CienciasmamalumbaAún no hay calificaciones
- Avance TICDocumento59 páginasAvance TICGeraldine FloresAún no hay calificaciones
- 2020 09 08 16 35 35 Valburqueque 03-09-Sistema-Nervioso2do-3erosec PDFDocumento20 páginas2020 09 08 16 35 35 Valburqueque 03-09-Sistema-Nervioso2do-3erosec PDFJunior Kevin Zavaleta ValverdeAún no hay calificaciones
- Areas Nasales TacDocumento9 páginasAreas Nasales TacporoheAún no hay calificaciones
- Análisis Película Yo Soy SamDocumento5 páginasAnálisis Película Yo Soy Samgalo Galo472Aún no hay calificaciones
- Un Viaje Hacia La VerdadDocumento2 páginasUn Viaje Hacia La VerdadAngela Correa100% (1)
- Transcripción y TraducciónDocumento6 páginasTranscripción y TraducciónJose Luis VallejosAún no hay calificaciones
- De: Mireia Nieto: ¿Qué Es y para Qué Sirve La Genealogía?Documento4 páginasDe: Mireia Nieto: ¿Qué Es y para Qué Sirve La Genealogía?Isaac VeraAún no hay calificaciones
- ELABORACION DE QUESO FRESCO Ing. FocDocumento11 páginasELABORACION DE QUESO FRESCO Ing. FocMp PatriciaAún no hay calificaciones
- MOSCADocumento19 páginasMOSCAMilagros MartinezAún no hay calificaciones
- 80 Preguntas Sobre Personalidad en Una Entrevista de TrabajoDocumento4 páginas80 Preguntas Sobre Personalidad en Una Entrevista de TrabajoAntonio Paulo Sepúlveda Muñoz100% (1)
- Orden de Los Componentes en Una NPTDocumento14 páginasOrden de Los Componentes en Una NPTluis fernando loaiza correaAún no hay calificaciones
- Gramineas PerennesDocumento85 páginasGramineas Perenneslillo777Aún no hay calificaciones
- Tarea Noemi (Autoguardado) NuevoDocumento8 páginasTarea Noemi (Autoguardado) Nuevodaniel david gonzalezAún no hay calificaciones
- MG Quat Sanitizer HDS Marzo 2019Documento5 páginasMG Quat Sanitizer HDS Marzo 2019GladyAún no hay calificaciones
- Pérez Pardo José Lucas. Viabilidad Del Digestato en Reactores de Vinazas para La Alimentación Animal.Documento84 páginasPérez Pardo José Lucas. Viabilidad Del Digestato en Reactores de Vinazas para La Alimentación Animal.Jose Lucas Perez Pardo100% (1)
- Pubalgia PDFDocumento6 páginasPubalgia PDFRobertto SanchezAún no hay calificaciones
- Desarrollo Embrionario de Peces AnualesDocumento1 páginaDesarrollo Embrionario de Peces Anualestheblack08Aún no hay calificaciones
- 1° Práctica (C2)Documento1 página1° Práctica (C2)cesar JuniorAún no hay calificaciones
- S2 Filogenia y OntogeniaDocumento48 páginasS2 Filogenia y OntogeniaJesus BabelicoAún no hay calificaciones