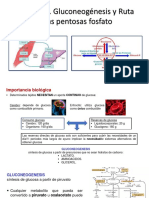
También podría gustarte
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- Ejercicio N°10 - Bioquímica - OCWDocumento4 páginasEjercicio N°10 - Bioquímica - OCWYan HernándezAún no hay calificaciones
- Mecanismo de TransducciónDocumento1 páginaMecanismo de TransducciónAbraham Landa100% (1)
- Respuestas Del Taller V - BCM 2021Documento23 páginasRespuestas Del Taller V - BCM 2021julicatu90Aún no hay calificaciones
- Preguntas GlucolisisDocumento14 páginasPreguntas GlucolisisAndy LucasAún no hay calificaciones
- GluconeogenesisDocumento24 páginasGluconeogenesisEloy Alfonso Flores CastilloAún no hay calificaciones
- Cap 8 GluconeogénsisDocumento29 páginasCap 8 Gluconeogénsissandily deysi pauccar panihuaraAún no hay calificaciones
- Apuntes de GluconeogénesisDocumento6 páginasApuntes de GluconeogénesisIsba Shadai Estrada GarciaAún no hay calificaciones
- Biosintesis y Degradacion Del GlucógenoDocumento31 páginasBiosintesis y Degradacion Del GlucógenoAaron QuispeAún no hay calificaciones
- Resumen 2do Parcial QuimicaDocumento28 páginasResumen 2do Parcial QuimicaFranco FilippoliAún no hay calificaciones
- Gluconeogénesis - Reacciones Enzimáticas - Dr. Alberto SanagustínDocumento5 páginasGluconeogénesis - Reacciones Enzimáticas - Dr. Alberto SanagustínJosmar Villano BonillaAún no hay calificaciones
- Tema 8 - Gluconeogenesis - RegulacionDocumento27 páginasTema 8 - Gluconeogenesis - RegulacionJoãoAún no hay calificaciones
- GluconeogénesisDocumento6 páginasGluconeogénesisJOSELY CARLOSAún no hay calificaciones
- Gluconeogenesis 2Documento73 páginasGluconeogenesis 2eduardoAún no hay calificaciones
- GluconeogenesisDocumento20 páginasGluconeogenesisdan100% (2)
- Seminario Gluconeogénesis RealizadoDocumento6 páginasSeminario Gluconeogénesis RealizadoLuz Jasmine FigueroaAún no hay calificaciones
- Wuolah-Free-Cuestiones de Exámenes PDFDocumento7 páginasWuolah-Free-Cuestiones de Exámenes PDFanon_373936375Aún no hay calificaciones
- Tarea1 Unidad4Documento12 páginasTarea1 Unidad4Abrahamalberto HernandezAún no hay calificaciones
- Gluconeogenesis PPT YesiDocumento20 páginasGluconeogenesis PPT YesiCesar Sarmiento PerezAún no hay calificaciones
- Ilovepdf MergedDocumento67 páginasIlovepdf MergedEl joven GilAún no hay calificaciones
- Apuntes BioqDocumento4 páginasApuntes BioqVanessaAún no hay calificaciones
- Gluconeogénesis, Glucogenogénesis y GlucogenólisisDocumento80 páginasGluconeogénesis, Glucogenogénesis y GlucogenólisisValdovinos KennethAún no hay calificaciones
- Solemnes BioquimicaDocumento7 páginasSolemnes BioquimicaSophieAún no hay calificaciones
- Bioquimica RepasoDocumento13 páginasBioquimica RepasoholaAún no hay calificaciones
- Tema 9 - Gluconeogénesis y Su RegulaciónDocumento13 páginasTema 9 - Gluconeogénesis y Su RegulaciónLauraAún no hay calificaciones
- SEMANA 8. Gluconeogénesis y Ruta de Las Pentosas FosfatoDocumento44 páginasSEMANA 8. Gluconeogénesis y Ruta de Las Pentosas FosfatoVIVIANA ZULEMA AZA�ERO MAGANAún no hay calificaciones
- GluconeogenesisDocumento37 páginasGluconeogenesisStefanny Gabriela100% (1)
- Gluconeogénesis - Ciclo de Las PentosasDocumento57 páginasGluconeogénesis - Ciclo de Las PentosasUSMP FN ARCHIVOSAún no hay calificaciones
- Gluconeogénesis, Glucogenóli Sis, Glucogénesis, Via de Las Pentosa FosfatoDocumento43 páginasGluconeogénesis, Glucogenóli Sis, Glucogénesis, Via de Las Pentosa FosfatoBertha GuivinAún no hay calificaciones
- OVA TEORIA - SEM 06 Glicólisis-CKDocumento38 páginasOVA TEORIA - SEM 06 Glicólisis-CKMarilyn TantaquispeAún no hay calificaciones
- UNIDAD IV B - Gluconeoge - Nesis - 21 Abril2016Documento30 páginasUNIDAD IV B - Gluconeoge - Nesis - 21 Abril2016Natalia Paz Solar MartinezAún no hay calificaciones
- Rutas Metabolicas CHOSDocumento51 páginasRutas Metabolicas CHOSAlfredo GonzalezAún no hay calificaciones
- Tema 3. Metabolismo de Carbohidratos. Gluconeogénesis.Documento8 páginasTema 3. Metabolismo de Carbohidratos. Gluconeogénesis.helloAún no hay calificaciones
- Cuestionario 2 - GluconeogénesisDocumento8 páginasCuestionario 2 - GluconeogénesisVanessa Lopez FAún no hay calificaciones
- Gluconeogenesis 1Documento34 páginasGluconeogenesis 1Ernesto GarciaAún no hay calificaciones
- TALLER#3Documento6 páginasTALLER#3Andy MoreiraAún no hay calificaciones
- 3.1 GluconeogensisDocumento18 páginas3.1 GluconeogensisAlfredo PalaciosAún no hay calificaciones
- Metabolismo de CarboidratosDocumento23 páginasMetabolismo de CarboidratosVerónica VelásquezAún no hay calificaciones
- Esquema de Catabolismo de Los CarbohidratosDocumento5 páginasEsquema de Catabolismo de Los CarbohidratosJuanPérezAún no hay calificaciones
- Resumen Metabolismo de Glúcidos, Regulación Metabolica, Metabolismo Glucogeno y Vias de Las Pentosas FosfatoDocumento28 páginasResumen Metabolismo de Glúcidos, Regulación Metabolica, Metabolismo Glucogeno y Vias de Las Pentosas FosfatocarolinaAún no hay calificaciones
- Gluconeogénesisglucogenólisisglucogénesisvia de Las Pentosa Fosfato 3Documento43 páginasGluconeogénesisglucogenólisisglucogénesisvia de Las Pentosa Fosfato 3Pilar Polo GonzalezAún no hay calificaciones
- Anabolismo de CarbohidratosDocumento21 páginasAnabolismo de Carbohidratosjessica benitez duarteAún no hay calificaciones
- Capítulos 18 Harper GlucólisisDocumento8 páginasCapítulos 18 Harper GlucólisisKensly GutierrezAún no hay calificaciones
- Bioquimica: GluconeogenesisDocumento39 páginasBioquimica: GluconeogenesisMedSchoolStuffAún no hay calificaciones
- Gluco Neo GenesisDocumento21 páginasGluco Neo GenesisFaiwon ParkAún no hay calificaciones
- Bioquímica Gluneogenesis y GlugenolisisDocumento104 páginasBioquímica Gluneogenesis y GlugenolisisDANNA VANESSA FLOREZ RUEDAAún no hay calificaciones
- Expo Bioquimica Metabolismo de Carbohidratos 2023Documento24 páginasExpo Bioquimica Metabolismo de Carbohidratos 2023Jhonny Alejandro Chadid BenítezAún no hay calificaciones
- Metabolismo de Los Hidratos de CarbonoDocumento4 páginasMetabolismo de Los Hidratos de CarbonoLara OviedoAún no hay calificaciones
- GlucólisisDocumento10 páginasGlucólisismaria moranAún no hay calificaciones
- Metabolismo de GlucogenoDocumento31 páginasMetabolismo de GlucogenoFelipe LavínAún no hay calificaciones
- Tema 7. Glucolisis - RegulacionDocumento28 páginasTema 7. Glucolisis - RegulacionJoãoAún no hay calificaciones
- Metabo C2Documento9 páginasMetabo C2Andrea MoralesAún no hay calificaciones
- Cuestionario Final. CocoDocumento17 páginasCuestionario Final. CocoAndrea MoralesAún no hay calificaciones
- Tem 6 BioqDocumento10 páginasTem 6 BioqAlexandra María Marín BernalAún no hay calificaciones
- Glucolisis y La Oxidacion de PiruvatoDocumento8 páginasGlucolisis y La Oxidacion de PiruvatoBrenda HAAún no hay calificaciones
- 4-UP 3 Glúcidos II (Con Audio)Documento66 páginas4-UP 3 Glúcidos II (Con Audio)tiziana.batistuttiAún no hay calificaciones
- Tema 8.metabolismo de Los Carbohidratos 12-11Documento34 páginasTema 8.metabolismo de Los Carbohidratos 12-11Yesica Alvis limaAún no hay calificaciones
- Segundo RepasoDocumento162 páginasSegundo Repasoevelyn regina martinez juarezAún no hay calificaciones
- Cuestionario ResueltoDocumento13 páginasCuestionario ResueltoJose Bastidas0% (1)
- Guia de Estudio de Metabolismo de CarbohidratosDocumento14 páginasGuia de Estudio de Metabolismo de CarbohidratosMelissa MartinezanguloAún no hay calificaciones
- Apuntes de EnzimasDocumento4 páginasApuntes de EnzimasSara Leiva LedezmaAún no hay calificaciones
- Curso Preuniversitario de Bioquímica 2020Documento3 páginasCurso Preuniversitario de Bioquímica 2020Dayi AmbrosiAún no hay calificaciones
- Resumen BioseñalizacionDocumento3 páginasResumen BioseñalizacionIsrael Fabian Valenzuela TeranAún no hay calificaciones
- 12 Biosíntesis y Almacenamiento de Ácidos GrasosDocumento3 páginas12 Biosíntesis y Almacenamiento de Ácidos Grasosjairo santiagoAún no hay calificaciones
- FOSFOLÍPIDOSDocumento2 páginasFOSFOLÍPIDOSRocio DLAún no hay calificaciones
- Tabla de Alimentos Ecuatorianos 1Documento12 páginasTabla de Alimentos Ecuatorianos 1Lili Moreta100% (1)
- Activdad. Introducción CarbohidratosDocumento2 páginasActivdad. Introducción CarbohidratosKelin Joselin Ceniceros SotoAún no hay calificaciones
- Bioquimica Del Ejercicio (073-102) .En - EsDocumento30 páginasBioquimica Del Ejercicio (073-102) .En - EsSel TMAún no hay calificaciones
- Moleculas y MacromoleculasDocumento38 páginasMoleculas y MacromoleculasProfesor Álvaro ProvosteAún no hay calificaciones
- ¿Qué Son Los CarbohidratosDocumento4 páginas¿Qué Son Los CarbohidratosAna CarrilloAún no hay calificaciones
- ACIDOS NUCLEÍCOS - PPT (7) .pptIIDocumento92 páginasACIDOS NUCLEÍCOS - PPT (7) .pptIIDaisy VanessaAún no hay calificaciones
- Tema 22-La TraducciónDocumento15 páginasTema 22-La TraducciónLauraAún no hay calificaciones
- Actividad N°08 ProteinasDocumento2 páginasActividad N°08 ProteinasSócrates TomazAún no hay calificaciones
- VITAMINASDocumento22 páginasVITAMINASÁngela DíazAún no hay calificaciones
- Cap. 03 Bases Moleculares Dr. Edson Ayala Quintanilla - 111518Documento27 páginasCap. 03 Bases Moleculares Dr. Edson Ayala Quintanilla - 111518Ghilmar Dennis Perez CasanovaAún no hay calificaciones
- Biologia 177-218 CiDocumento40 páginasBiologia 177-218 CiAzulé HerediaAún no hay calificaciones
- Bioquimica de La InsulinaDocumento2 páginasBioquimica de La Insulinayocelin aguilarAún no hay calificaciones
- Diapositivas Unidad 2 Bioquimica Grecia Peralta D-01Documento10 páginasDiapositivas Unidad 2 Bioquimica Grecia Peralta D-01GreciaAún no hay calificaciones
- EzimasDocumento6 páginasEzimasdifat franciscoAún no hay calificaciones
- Bio Qui MicaDocumento4 páginasBio Qui MicaSergio Velasquez Torres100% (1)
- La Información HereditariaDocumento17 páginasLa Información HereditariaErika KelsyAún no hay calificaciones
- 1 Cátalogo de Tarjetas 1626277934829629Documento17 páginas1 Cátalogo de Tarjetas 1626277934829629Diana Gabriela NinaAún no hay calificaciones
- Qué Es El ATPDocumento3 páginasQué Es El ATPCharlie MartinAún no hay calificaciones
- Taller Proteinas y Enzimas PDFDocumento3 páginasTaller Proteinas y Enzimas PDFCyberlosophy BlogAún no hay calificaciones
- Dogma CelularDocumento50 páginasDogma CelularAllison ParraAún no hay calificaciones
- Receptores CelularesDocumento2 páginasReceptores CelularesALLISSON YACHIMBAAún no hay calificaciones
- Practica Identificación Desnaturalización y Acción Enzimatica de Las ProteínasDocumento5 páginasPractica Identificación Desnaturalización y Acción Enzimatica de Las ProteínasSamantha Morales MartinezAún no hay calificaciones
- Digestión de ProteínasDocumento9 páginasDigestión de Proteínasmario villaAún no hay calificaciones
- Taller de Recuperación de Primer Periodo Grado 9°Documento5 páginasTaller de Recuperación de Primer Periodo Grado 9°sandra maldonadoAún no hay calificaciones