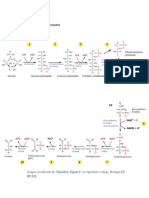
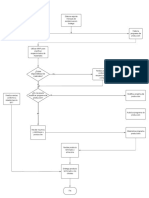
También podría gustarte
- DISLIPIDEMIADocumento5 páginasDISLIPIDEMIAMiguel Ángel Jesús Lavarello BermejoAún no hay calificaciones
- Regulación de Las Funciones Corporales y HomeostasisDocumento13 páginasRegulación de Las Funciones Corporales y HomeostasisCristian FloresAún no hay calificaciones
- Guia de Integracion Metabolica CompletaDocumento19 páginasGuia de Integracion Metabolica CompletaBrianAún no hay calificaciones
- Discusion 3Documento17 páginasDiscusion 3Milver EscalanteAún no hay calificaciones
- CK (Spinreact)Documento1 páginaCK (Spinreact)Arfaxad CercleAún no hay calificaciones
- DislipidemiasDocumento8 páginasDislipidemiasSCOMP AEFMUAC100% (1)
- Bioquimica Practica #9 Trigliceridos IiDocumento12 páginasBioquimica Practica #9 Trigliceridos Iifrank ganz bravoAún no hay calificaciones
- Enzimas de escape clínicasDocumento43 páginasEnzimas de escape clínicasRosas Ramírez Omar0% (1)
- Sind. Hipo e Hipersecreción de InsulinaDocumento38 páginasSind. Hipo e Hipersecreción de Insulinahemer666100% (5)
- Dislipidemia en adulto mayorDocumento7 páginasDislipidemia en adulto mayormanuel nampulaAún no hay calificaciones
- DESLIPIDEMIADocumento19 páginasDESLIPIDEMIAPiero Paolo100% (1)
- Cetoacidosis Diabética: Complicación Metabólica Aguda en Pacientes con DMDocumento5 páginasCetoacidosis Diabética: Complicación Metabólica Aguda en Pacientes con DMLina OrjuelaAún no hay calificaciones
- DislipidemiasDocumento41 páginasDislipidemiasDiego CidAún no hay calificaciones
- Acidosis Metabolica en UciDocumento34 páginasAcidosis Metabolica en UciArturo1985A100% (1)
- Isoenzimas de La AmilasaDocumento4 páginasIsoenzimas de La AmilasaAlbertoAún no hay calificaciones
- Metabolismo de Compuestos Nitrogenados 1Documento41 páginasMetabolismo de Compuestos Nitrogenados 1juanAún no hay calificaciones
- Presentación Intoxicación Por Yuca AmargaDocumento19 páginasPresentación Intoxicación Por Yuca AmargaAlbert RattiaAún no hay calificaciones
- Práctica 5 - 1 - LìpidosDocumento9 páginasPráctica 5 - 1 - LìpidosIvan NuñezAún no hay calificaciones
- Dislipidemias y ArteriosclerosisDocumento20 páginasDislipidemias y ArteriosclerosisClaudia Patricia Ballen GarzonAún no hay calificaciones
- Caso Clinico DHCDocumento4 páginasCaso Clinico DHCKarito FigueroaAún no hay calificaciones
- DM Tipo 3Documento8 páginasDM Tipo 3Urías JaraAún no hay calificaciones
- Sangrado Del Primer TrimestreDocumento42 páginasSangrado Del Primer TrimestreveguetaAún no hay calificaciones
- LipoproteinasDocumento6 páginasLipoproteinasPedro Guillermo BarretoAún no hay calificaciones
- Preguntas para El ExamenDocumento4 páginasPreguntas para El ExamenElsa Santos tocasAún no hay calificaciones
- Enfermedad de TangierDocumento4 páginasEnfermedad de TangierGénesis Alejandra Tapia ChamorroAún no hay calificaciones
- 3 LipoproteinasDocumento46 páginas3 Lipoproteinasjackriden100% (1)
- Teoría Perfil de Lípidos Otoño 2021Documento18 páginasTeoría Perfil de Lípidos Otoño 2021Josue Azcue100% (1)
- Hemoglobina FetalDocumento3 páginasHemoglobina FetalPayola TitoAún no hay calificaciones
- Cuestionario Fisiopatología - Diabetes MellitusDocumento7 páginasCuestionario Fisiopatología - Diabetes MellitusCarla Marliz Ore Jaime100% (1)
- Práctica 1 Química SanguíneaDocumento14 páginasPráctica 1 Química SanguíneaSandy MoralesAún no hay calificaciones
- Ensayo NUEVE (Proteínas Totales Globulinas y Albúminas)Documento6 páginasEnsayo NUEVE (Proteínas Totales Globulinas y Albúminas)Cbz Sebastian Zermeño BrionesAún no hay calificaciones
- Albumina 2 1Documento2 páginasAlbumina 2 1Yadira Hernandez Ramirez100% (1)
- Bioquimica 2 ParcialDocumento18 páginasBioquimica 2 ParcialAngelaMedinaDAún no hay calificaciones
- Síntesis de ácidos grasos y beta-reducción en la lipogénesisDocumento23 páginasSíntesis de ácidos grasos y beta-reducción en la lipogénesisAdylene Montes MAún no hay calificaciones
- HALOPERIDOLDocumento15 páginasHALOPERIDOLRivaldo VergaraAún no hay calificaciones
- Transporte de GlucosaDocumento9 páginasTransporte de GlucosaRafael GaytánAún no hay calificaciones
- Terapias Hipoglucemiantes e Insuficiencia Cardíaca en La Diabetes Mellitus Tipo 2-1Documento12 páginasTerapias Hipoglucemiantes e Insuficiencia Cardíaca en La Diabetes Mellitus Tipo 2-1Iris Gutiérrez100% (1)
- Taller de LipidosDocumento4 páginasTaller de LipidosTatiana RodriguezAún no hay calificaciones
- Clase: Nutrición y CancerDocumento42 páginasClase: Nutrición y CancerLuna SheccidAún no hay calificaciones
- Lipoproteínas plasmáticas guía de estudioDocumento5 páginasLipoproteínas plasmáticas guía de estudiochristian100% (1)
- Determinación de triglicéridos en sueroDocumento2 páginasDeterminación de triglicéridos en sueroNilbia GarciaAún no hay calificaciones
- Emergencias Metabólicas Agudas en DiabetesDocumento38 páginasEmergencias Metabólicas Agudas en DiabetesRomulo Alegre LeónAún no hay calificaciones
- Dislipidemia InformeDocumento17 páginasDislipidemia InformeFrancisco Javier Madrid RuizAún no hay calificaciones
- Disbetalipoproteinemia FamiliarDocumento3 páginasDisbetalipoproteinemia Familiarapi-3697492Aún no hay calificaciones
- AlbuminaDocumento9 páginasAlbuminaJosue Santiago LopezAún no hay calificaciones
- Laboratorio Tabla de Valores Normales PDFDocumento5 páginasLaboratorio Tabla de Valores Normales PDFLE JuarezAún no hay calificaciones
- Discusion de Grupo No 4Documento22 páginasDiscusion de Grupo No 4JenicitaGracias100% (4)
- Caso Clinico DiabetesDocumento2 páginasCaso Clinico DiabetesAndrea Jáuregui100% (1)
- Exposicion de Bioquimica Alterac. Metabolismo LípidosDocumento30 páginasExposicion de Bioquimica Alterac. Metabolismo Lípidosjulissa chacpiAún no hay calificaciones
- GlicopeptidosDocumento17 páginasGlicopeptidosapi-3750373100% (1)
- Encefalopatía HepáticaDocumento51 páginasEncefalopatía HepáticaPaulichie100% (11)
- Determinación de Colesterol TotalDocumento3 páginasDeterminación de Colesterol TotalAna Teresa Luna OliveraAún no hay calificaciones
- Bioquimica de La Diabetes 2Documento16 páginasBioquimica de La Diabetes 2AdonayDecoraciones50% (2)
- GluconeogenesisDocumento2 páginasGluconeogenesisAnnyVal RMAún no hay calificaciones
- Gluconeogenesis 2Documento73 páginasGluconeogenesis 2eduardoAún no hay calificaciones
- Bioquimica RepasoDocumento13 páginasBioquimica RepasoholaAún no hay calificaciones
- GluconeogénesisDocumento16 páginasGluconeogénesisNATALIA DANIELA VILLADIEGO BERROCALAún no hay calificaciones
- GLUCONEOGÉNESISDocumento14 páginasGLUCONEOGÉNESISNicoleAún no hay calificaciones
- Metabolismo de los hidratos de carbonoDocumento28 páginasMetabolismo de los hidratos de carbonoFranco FilippoliAún no hay calificaciones
- GLUCONEOGÉNESISDocumento22 páginasGLUCONEOGÉNESISyuri yuriAún no hay calificaciones
- Enzimas Plasmáticas de Interés ClínicoDocumento8 páginasEnzimas Plasmáticas de Interés ClínicoValdovinos KennethAún no hay calificaciones
- CarbohidratosDocumento13 páginasCarbohidratosValdovinos KennethAún no hay calificaciones
- Aspectos Químicos de La VidaDocumento4 páginasAspectos Químicos de La VidaValdovinos KennethAún no hay calificaciones
- Tarea 1. GlucólisisDocumento8 páginasTarea 1. GlucólisisValdovinos KennethAún no hay calificaciones
- Isomerà ADocumento45 páginasIsomerà AValdovinos KennethAún no hay calificaciones
- Organizacià N Quã - Mica de La Cã©lula.Documento98 páginasOrganizacià N Quã - Mica de La Cã©lula.Valdovinos KennethAún no hay calificaciones
- Distancia Acarreo EconomicoDocumento7 páginasDistancia Acarreo EconomiconinoAún no hay calificaciones
- Actividad 12 Fundamentos y Administracion de ComprasDocumento3 páginasActividad 12 Fundamentos y Administracion de ComprasAngie GuerreroAún no hay calificaciones
- Curso Ruso Intermedio AvanzadoDocumento10 páginasCurso Ruso Intermedio Avanzado012345550% (2)
- Microcurriculo FichaDocumento2 páginasMicrocurriculo FichaYina Fabiola Castro OchoaAún no hay calificaciones
- Ficha Salud 2022Documento1 páginaFicha Salud 2022Paula Vanessa Orjuela CiprianAún no hay calificaciones
- Fargo Dtc1250eDocumento82 páginasFargo Dtc1250eGeorge Orange0% (1)
- S3 - A1 - Artículo "El Reconocimiento Del Líder Educativo Como Agente de CambioDocumento2 páginasS3 - A1 - Artículo "El Reconocimiento Del Líder Educativo Como Agente de CambioTS. RODOLFO SÁNCHEZ TABOADA0% (1)
- Tarea I Planificacion y Gestion AulicaDocumento4 páginasTarea I Planificacion y Gestion AulicaAnonymous lQmo2HnCAún no hay calificaciones
- Mapa ProfesiogramaDocumento1 páginaMapa ProfesiogramadiogoAún no hay calificaciones
- Proforma de libros para la Universidad Nacional Santiago Antunez de MayoloDocumento38 páginasProforma de libros para la Universidad Nacional Santiago Antunez de MayoloRovert Sacramento AlvinagortaAún no hay calificaciones
- Hipias MenorDocumento12 páginasHipias MenorangieAún no hay calificaciones
- Programa de trabajo seguro en alturasDocumento50 páginasPrograma de trabajo seguro en alturasOmar Montoya33% (3)
- Diagramas de FlujoDocumento1 páginaDiagramas de Flujoelias anculleAún no hay calificaciones
- Laboratorio 6Documento8 páginasLaboratorio 6Salua zazir Carmona CarrmonaAún no hay calificaciones
- Semana 5Documento5 páginasSemana 5nelson huaytaAún no hay calificaciones
- Terapia EstrategicaDocumento24 páginasTerapia EstrategicaItalo B. Lecca100% (1)
- Unidad 6 Seguridad y Salud LaboralDocumento13 páginasUnidad 6 Seguridad y Salud LaboralLazaaAún no hay calificaciones
- Las Nuevas Teorias Del ConocimientoDocumento13 páginasLas Nuevas Teorias Del ConocimientoOrlando Valadez0% (1)
- LEY 1012 de Arancel de Honorarios de ArquitectosDocumento7 páginasLEY 1012 de Arancel de Honorarios de Arquitectosdanves007100% (1)
- Tarea 2. - Sistemas de Produccion.Documento7 páginasTarea 2. - Sistemas de Produccion.JOSE JESUS MARTINEZ ACOSTAAún no hay calificaciones
- DiagnosticoyclasificacionPsiquiatria PDFDocumento11 páginasDiagnosticoyclasificacionPsiquiatria PDFMariela CespedesAún no hay calificaciones
- Temporizadores ENYA Serie - Retardos, Impulsos y Ciclos SimétricosDocumento3 páginasTemporizadores ENYA Serie - Retardos, Impulsos y Ciclos SimétricosClaudio Patricio Zarate VidalAún no hay calificaciones
- La Transformación Chamanica Del Hombre en JaguarDocumento15 páginasLa Transformación Chamanica Del Hombre en JaguarFredy Segura100% (1)
- AntropometriaDocumento27 páginasAntropometriaAdriana Gómez MorenoAún no hay calificaciones
- Cómo Realizar Ensayo Densidad en El Sitio Con El Cono de ArenaDocumento21 páginasCómo Realizar Ensayo Densidad en El Sitio Con El Cono de ArenaMarckTkmMaznaAún no hay calificaciones
- Actividades de Informatica para NinosDocumento11 páginasActividades de Informatica para NinosAlejandra Morales80% (5)
- Aguilar VillanuevaDocumento2 páginasAguilar VillanuevaJoaquin AscaniAún no hay calificaciones
- Analisis Granulometrico InformeDocumento6 páginasAnalisis Granulometrico InformeAldo Sabino Flores Sapacayo100% (2)
- Guía # 1 Introducción Al Estudio de La ÉticaDocumento15 páginasGuía # 1 Introducción Al Estudio de La Éticairma Areli GalloAún no hay calificaciones
- CiudadrepresentadaDocumento13 páginasCiudadrepresentadaJulio Motörhead GuascaAún no hay calificaciones