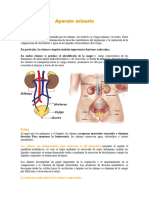
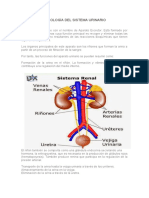
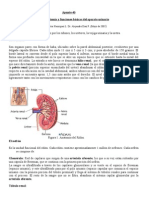
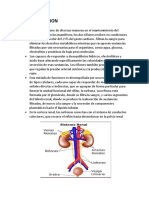
También podría gustarte
- Aparato UrinarioDocumento19 páginasAparato UrinarioAnaAún no hay calificaciones
- Sistema UrinarioDocumento20 páginasSistema UrinarioPatricia PetersAún no hay calificaciones
- Resumen Sistema RenalDocumento8 páginasResumen Sistema RenalXochitl C. Palomec100% (3)
- Edoc - Tips - Regulacion de Los Liquidos Extracelulares y de La Sangre Por LosDocumento7 páginasEdoc - Tips - Regulacion de Los Liquidos Extracelulares y de La Sangre Por LosAbril QuinnAún no hay calificaciones
- Anatomía y Fisiología RenalDocumento10 páginasAnatomía y Fisiología RenalJoan william Salazar paguasiAún no hay calificaciones
- 3° - Guía de Estudio - Anatomía y Fisiología RenalDocumento12 páginas3° - Guía de Estudio - Anatomía y Fisiología RenalMiguel CandiaAún no hay calificaciones
- Sistema RenalDocumento7 páginasSistema RenalHeydi Cardeno PAún no hay calificaciones
- Exposicion BiologiaDocumento3 páginasExposicion BiologiaRoberto Carlos Correa AcuñaAún no hay calificaciones
- 6 Fisiología RenalDocumento15 páginas6 Fisiología RenalJasna ZovakAún no hay calificaciones
- Clase 1 y 2Documento43 páginasClase 1 y 2studyshop29Aún no hay calificaciones
- H. Sistema UrinarioDocumento15 páginasH. Sistema UrinarioLisbeth FigueroaAún no hay calificaciones
- Aparato Urinario Anatomia y FisiologiaDocumento11 páginasAparato Urinario Anatomia y Fisiologiayordane quispe delgadoAún no hay calificaciones
- 8 Fisiologia RenalDocumento34 páginas8 Fisiologia RenalINGO MANUEL RIECKHOFF SANCHEZAún no hay calificaciones
- Manual de EgoDocumento47 páginasManual de EgoRosenda DLAún no hay calificaciones
- Sistema UrinarioDocumento5 páginasSistema UrinarioalexiaguzzmanAún no hay calificaciones
- Renal HidrosalinoDocumento37 páginasRenal HidrosalinoViviana Hernandez QuijanoAún no hay calificaciones
- Cuestionario Del Sistema UrinarioDocumento6 páginasCuestionario Del Sistema UrinarioBurgos JhoanaAún no hay calificaciones
- Apuntes NefrologiaDocumento12 páginasApuntes NefrologiaVale ReynosoAún no hay calificaciones
- Anatomía y Fisiología Del Sistema UrinarioDocumento7 páginasAnatomía y Fisiología Del Sistema UrinarioMarcos LeonAún no hay calificaciones
- Fisiologia RenalDocumento47 páginasFisiologia RenalEliezer RiveroAún no hay calificaciones
- Aparato Urinario Histologia Grupo 5Documento4 páginasAparato Urinario Histologia Grupo 5Norka Mirella Uchamaco HuancaAún no hay calificaciones
- Anatomia y Fisiologia RenalDocumento48 páginasAnatomia y Fisiologia RenalAhuitzolt BravoAún no hay calificaciones
- Histologia Del Sistema Urinario ActualDocumento32 páginasHistologia Del Sistema Urinario ActualAshley MaradiagaAún no hay calificaciones
- HistologíaDocumento7 páginasHistologíaFrancelys MendozaAún no hay calificaciones
- Anatomia y Fisiologia RenalDocumento12 páginasAnatomia y Fisiologia RenalCarlos Gonzalez AlemanAún no hay calificaciones
- PROPIEDADES DEL RIÑON, FILTRACION GLOMERULAR. (Autoguardado)Documento30 páginasPROPIEDADES DEL RIÑON, FILTRACION GLOMERULAR. (Autoguardado)ab21023Aún no hay calificaciones
- HistoDocumento6 páginasHistoPatricia Salazar ZambranoAún no hay calificaciones
- ClasesDocumento37 páginasClasesJhoana velascoAún no hay calificaciones
- Apuntes NefronaDocumento8 páginasApuntes NefronaNinfa Esther Montejo de la CruzAún no hay calificaciones
- Sistema RenalDocumento102 páginasSistema RenalAdry FaryAún no hay calificaciones
- HACER CLICK AQUÍ PARA DESCARGAR Guia 8° Sistema Renal 2012 Prof. Arturo Riveros PDFDocumento5 páginasHACER CLICK AQUÍ PARA DESCARGAR Guia 8° Sistema Renal 2012 Prof. Arturo Riveros PDFDamaris Betsabe Arancibia MolinaAún no hay calificaciones
- Embrio 2 c3 S.renal CompletoDocumento18 páginasEmbrio 2 c3 S.renal CompletoJENNIFER DANIELA MARTINEZ LOPEZAún no hay calificaciones
- Apunte46 Anatomía y Funciones Básicas Del Aparato UrinarioDocumento6 páginasApunte46 Anatomía y Funciones Básicas Del Aparato UrinarioDANIEL ALEJANDROAún no hay calificaciones
- Sistema RenalDocumento31 páginasSistema Renaljuan pedro100% (2)
- Funciones de Los RiñonesDocumento12 páginasFunciones de Los RiñonesMario Antonio Romero AgüeroAún no hay calificaciones
- Sistema Urinario 2021Documento9 páginasSistema Urinario 2021Cristofer Oliver Peñaloza YujraAún no hay calificaciones
- 11 Aparato Urinario - Apunte de CátedraDocumento9 páginas11 Aparato Urinario - Apunte de CátedraLuz OlivaAún no hay calificaciones
- Funcion RenalDocumento10 páginasFuncion RenalNiicolePinedaAún no hay calificaciones
- Fisiología Resumen Medicina Segundo Año. Sistema Renal y Digestivo Universidad de SantiagoDocumento40 páginasFisiología Resumen Medicina Segundo Año. Sistema Renal y Digestivo Universidad de SantiagoClaudio Alexis CifuentesAún no hay calificaciones
- Resumen Sistema UrinarioDocumento9 páginasResumen Sistema UrinarioMarina Morales100% (1)
- Bibliografía Recomendad1Documento25 páginasBibliografía Recomendad1Iglesia Sendero De SalvacionAún no hay calificaciones
- Histologia Sistema UrinarioDocumento4 páginasHistologia Sistema Urinariocheveloso100% (2)
- Sistema UrinarioDocumento8 páginasSistema Urinariomichelle urrutiaAún no hay calificaciones
- RESUMEN APARATO URINARIO - TortoraDocumento5 páginasRESUMEN APARATO URINARIO - TortoraFerminAún no hay calificaciones
- Aparato UrinarioDocumento64 páginasAparato UrinarioSalvador BARRON MELOAún no hay calificaciones
- Fisiologia RenalDocumento11 páginasFisiologia RenalJonathan ArmijosAún no hay calificaciones
- Elementos de La Función RenalDocumento8 páginasElementos de La Función RenalFlorencia BarsiAún no hay calificaciones
- Tema 8 ArregloDocumento30 páginasTema 8 ArregloGonzalo Sánchez DiéguezAún no hay calificaciones
- Manual de E.G.O.Documento50 páginasManual de E.G.O.Nino Flores100% (1)
- MORFOFISIOLOGÍA RENAL (1) - ComprimidoDocumento22 páginasMORFOFISIOLOGÍA RENAL (1) - ComprimidoAdolfo Fuentes SánchezAún no hay calificaciones
- Tarea Del Primer Parcial de GenitourinarioDocumento9 páginasTarea Del Primer Parcial de GenitourinarioRetdiniAún no hay calificaciones
- Disección Del RiñonDocumento13 páginasDisección Del RiñonDanny Troya GomezAún no hay calificaciones
- Aparato UrinarioDocumento5 páginasAparato UrinarioDaniel MéndezAún no hay calificaciones
- Notitas RenalDocumento18 páginasNotitas RenalEmilia BenitezAún no hay calificaciones
- Apunte FisiologiaDocumento15 páginasApunte FisiologiaTeresaAún no hay calificaciones
- Anatomía y Fisiología RenalDocumento51 páginasAnatomía y Fisiología RenalMario Nieto'80% (5)
- Fisiologia RenalDocumento21 páginasFisiologia RenalDayana Ruiz HerreraAún no hay calificaciones
- Renal Fisiología Humana IDocumento81 páginasRenal Fisiología Humana INora El KhamlichiAún no hay calificaciones
- Introducción Básica a la Microcirculación de la Sangre: La Siguiente Generación en Cuidados de la SaludDe EverandIntroducción Básica a la Microcirculación de la Sangre: La Siguiente Generación en Cuidados de la SaludAún no hay calificaciones
- Coagulación Para Todos: Medicina Para TodosDe EverandCoagulación Para Todos: Medicina Para TodosCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Practica No.4 A 8Documento3 páginasPractica No.4 A 8Evelina CidAún no hay calificaciones
- Historia de PSI en RDDocumento20 páginasHistoria de PSI en RDAbelAún no hay calificaciones
- Apuntes de AnatomíaDocumento1 páginaApuntes de AnatomíaEvelina CidAún no hay calificaciones
- Quiz HistDocumento1 páginaQuiz HistEvelina CidAún no hay calificaciones
- Aparato RespiratorioDocumento84 páginasAparato RespiratorioRosangelyy RosarioAún no hay calificaciones
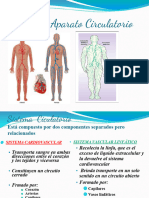
- Aparato CirculatorioDocumento97 páginasAparato CirculatorioEvelina CidAún no hay calificaciones
- PielDocumento81 páginasPielEvelina CidAún no hay calificaciones
- Microbiología. Personajes de La MicrobiologíaDocumento3 páginasMicrobiología. Personajes de La MicrobiologíaEvelina CidAún no hay calificaciones
- Cavidad Oral y Glandulas SalivalesDocumento77 páginasCavidad Oral y Glandulas SalivalesEvelina CidAún no hay calificaciones
- Fluidos ArcillaDocumento27 páginasFluidos ArcillaJobita JMAún no hay calificaciones
- Corrientes GalvanicasDocumento24 páginasCorrientes GalvanicasLiliana LeguizamonAún no hay calificaciones
- Lab 01 - Fisicoquímica-3Documento10 páginasLab 01 - Fisicoquímica-3Maricielo CerroAún no hay calificaciones
- Examen Final Quimica AplicadaDocumento18 páginasExamen Final Quimica AplicadaMarcelo FrancoAún no hay calificaciones
- DIRTEX Catalogo General (Triptico) 2020 FBBDocumento6 páginasDIRTEX Catalogo General (Triptico) 2020 FBBLuis RetamozoAún no hay calificaciones
- 2023-Tema 04-SolucionesDocumento45 páginas2023-Tema 04-SolucionesCine TVAún no hay calificaciones
- Glucósidos Cardiotónicos y Saponinas2Documento48 páginasGlucósidos Cardiotónicos y Saponinas2Anderson FernandezAún no hay calificaciones
- Práctica #03 Separacion de MezclasDocumento4 páginasPráctica #03 Separacion de MezclasFIORELLA VANESSA GÜERE SALAZARAún no hay calificaciones
- Anexo 4 Ficha Tecnica Gestion Del BiogasDocumento19 páginasAnexo 4 Ficha Tecnica Gestion Del BiogasDiego SuarezAún no hay calificaciones
- Marco TeóricoDocumento12 páginasMarco TeóricoAljhailey D. Sifontes V.Aún no hay calificaciones
- Practica 1Documento3 páginasPractica 1Rafael CastilloAún no hay calificaciones
- Clase - 4 MUROS - y - TABIQUES PDFDocumento150 páginasClase - 4 MUROS - y - TABIQUES PDFEmilio Guerra U.100% (1)
- Guía de Trabajo, La Tabla PeriódicaDocumento4 páginasGuía de Trabajo, La Tabla Periódicajose cortez florezAún no hay calificaciones
- Practica de Laboratorio N°3 Volumenes Molares Parciales 1. ObjetivosDocumento4 páginasPractica de Laboratorio N°3 Volumenes Molares Parciales 1. ObjetivosJohnjairo Edu Romero EspinoAún no hay calificaciones
- IESTec Quimica 2021Documento37 páginasIESTec Quimica 2021ANIBAL VILLALBAAún no hay calificaciones
- Antoine Lavoiser Biografia y AportacionesDocumento4 páginasAntoine Lavoiser Biografia y AportacionesxjtgzAún no hay calificaciones
- DEOeditmonografiaDocumento73 páginasDEOeditmonografiaGuilhermino JosenoronhacardosoAún no hay calificaciones
- Mapa BIOLOGIADocumento5 páginasMapa BIOLOGIADavid Ayala SaavedraAún no hay calificaciones
- Practica #10 ANHIDRASA CARBÓNICA 1Documento7 páginasPractica #10 ANHIDRASA CARBÓNICA 1alvarod_61Aún no hay calificaciones
- Método Integral para Obtener El Orden y La Constante de VelocidadDocumento4 páginasMétodo Integral para Obtener El Orden y La Constante de Velocidadivan lopezAún no hay calificaciones
- HafnioDocumento3 páginasHafnioVictorAún no hay calificaciones
- Quimica OrganicaDocumento15 páginasQuimica OrganicaNadim SalomonAún no hay calificaciones
- Quimica 1ro 4Documento2 páginasQuimica 1ro 4poli2254Aún no hay calificaciones
- Reporte Sesion A AvanceDocumento23 páginasReporte Sesion A AvanceKimberly G. TreviñoAún no hay calificaciones
- Clase 11 30 - 03 - 22Documento29 páginasClase 11 30 - 03 - 22andres guerreroAún no hay calificaciones
- Practica de Laboratorio FotosíntesisDocumento7 páginasPractica de Laboratorio FotosíntesisMadeleineAún no hay calificaciones
- Fundición (Ensayo A Fuego)Documento7 páginasFundición (Ensayo A Fuego)Alberto MonroyAún no hay calificaciones
- Capitulo 25 - 2501Documento3 páginasCapitulo 25 - 2501aaguneraAún no hay calificaciones
- Tema 13 AnabolismoDocumento3 páginasTema 13 AnabolismoMara NúñezAún no hay calificaciones
- MedicinaDocumento34 páginasMedicinaFrancisco BenitesAún no hay calificaciones