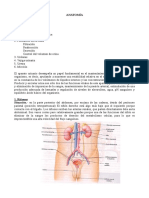
También podría gustarte
- Estructura y Funcion de La Esfera Urogenital IDocumento68 páginasEstructura y Funcion de La Esfera Urogenital IArlynes AriasAún no hay calificaciones
- Fisiologia Del RiñonDocumento16 páginasFisiologia Del RiñonEmily AsencioAún no hay calificaciones
- Bibliografía Recomendad1Documento25 páginasBibliografía Recomendad1Iglesia Sendero De SalvacionAún no hay calificaciones
- FISIOLOGIA RENAL - Morf.TeoDocumento4 páginasFISIOLOGIA RENAL - Morf.Teoyar sanadriAún no hay calificaciones
- FISIOLOGIA RENALDocumento21 páginasFISIOLOGIA RENALDayana Ruiz HerreraAún no hay calificaciones
- Fisiologia RenalDocumento13 páginasFisiologia RenalCarlos AquinoAún no hay calificaciones
- Sistema RenalDocumento14 páginasSistema RenalJennifer CalderónAún no hay calificaciones
- Fisiología RenalDocumento20 páginasFisiología RenalAngel GUAún no hay calificaciones
- Anatomia y Fisiologia Del Sistema ExcretorDocumento19 páginasAnatomia y Fisiologia Del Sistema ExcretorHernan MezaAún no hay calificaciones
- Estructura y Funcion de La Esfera UrogenitalDocumento65 páginasEstructura y Funcion de La Esfera UrogenitalArlynes AriasAún no hay calificaciones
- Anatomia y Fisiologia Del Sistema ExcretorDocumento17 páginasAnatomia y Fisiologia Del Sistema ExcretorRoberto Gustavo LlanesAún no hay calificaciones
- Nefrologia Dia 335 PDFDocumento23 páginasNefrologia Dia 335 PDFDannyMichelleAún no hay calificaciones
- Resumen Fisio RenalDocumento11 páginasResumen Fisio RenalNayla celmary González cunemoAún no hay calificaciones
- 7 Sistema Excretor - ResumenDocumento5 páginas7 Sistema Excretor - ResumenJasna ZovakAún no hay calificaciones
- Semana 14 - Fisiologia RenalDocumento62 páginasSemana 14 - Fisiologia RenalvivianajulonAún no hay calificaciones
- Funciones riñonesDocumento12 páginasFunciones riñonesEveline Pulinario LorenzoAún no hay calificaciones
- Nefrologia Dia 335Documento32 páginasNefrologia Dia 335Nadiia ValdeAún no hay calificaciones
- De Que Esta Compuesto El RiñonDocumento3 páginasDe Que Esta Compuesto El RiñonAlex Quintana0% (1)
- Fisiología RenalDocumento19 páginasFisiología Renaljuan pablo ospinaAún no hay calificaciones
- Resumen Sistema UrinarioDocumento9 páginasResumen Sistema UrinarioMarina Morales100% (1)
- Anatomia y Fisiologia de Los RiñonesDocumento17 páginasAnatomia y Fisiologia de Los RiñonesArturo Merida RodriguezAún no hay calificaciones
- Fisiologia RenalDocumento29 páginasFisiologia RenalLupe Torres TarazonaAún no hay calificaciones
- Sistema RenalDocumento31 páginasSistema Renaljuan pedro100% (2)
- Litiasis UrinariaDocumento9 páginasLitiasis UrinariaAdriana BenavidesAún no hay calificaciones
- UNIDAD 7 APARATO URINARIO ANATOMOFISIOLOGIA (1)Documento3 páginasUNIDAD 7 APARATO URINARIO ANATOMOFISIOLOGIA (1)Patricia SalgadoAún no hay calificaciones
- Sistema Urinario - BrendaDocumento40 páginasSistema Urinario - BrendaBrenda TrujilloAún no hay calificaciones
- Sistema Nefrourinario 2010Documento15 páginasSistema Nefrourinario 2010Nibaldo SaavedraAún no hay calificaciones
- ClasesDocumento37 páginasClasesJhoana velascoAún no hay calificaciones
- Presentación de Sistema Renal Genitourinario y HematopoyéticoDocumento289 páginasPresentación de Sistema Renal Genitourinario y Hematopoyéticopriscila monterrosaAún no hay calificaciones
- Glomerulonefritis - LoreDocumento18 páginasGlomerulonefritis - LoreAnto FrancoAún no hay calificaciones
- Función renal y anatomía básicaDocumento52 páginasFunción renal y anatomía básicaBrayan BarajasAún no hay calificaciones
- Anatomía y Fisiología RenalDocumento10 páginasAnatomía y Fisiología RenalJoan william Salazar paguasiAún no hay calificaciones
- Resumen Aparato UrinarioDocumento9 páginasResumen Aparato UrinarioFlorencia Ferraro ReyAún no hay calificaciones
- Función RenalDocumento8 páginasFunción RenalMayte TaquichiriAún no hay calificaciones
- Anatomia y Fisiologia RenalDocumento12 páginasAnatomia y Fisiologia RenalCarlos Gonzalez AlemanAún no hay calificaciones
- Sistema RenalDocumento10 páginasSistema RenalYINA MARCELA RUIZ LOMBANAAún no hay calificaciones
- Glomerulonefritis - EscritoDocumento19 páginasGlomerulonefritis - EscritoAnto FrancoAún no hay calificaciones
- RIÑONES. Seminario Fisiopatologia I. COMPLETOODocumento16 páginasRIÑONES. Seminario Fisiopatologia I. COMPLETOOtefa2602Aún no hay calificaciones
- Guia Sistema RenalDocumento5 páginasGuia Sistema RenalDavid FloresAún no hay calificaciones
- T 14 Aparato UrinarioDocumento7 páginasT 14 Aparato Urinarioapi-245849830Aún no hay calificaciones
- Renal Fisiología Humana IDocumento81 páginasRenal Fisiología Humana INora El KhamlichiAún no hay calificaciones
- Resumen AnatomoII-1Documento37 páginasResumen AnatomoII-1solicami yaAún no hay calificaciones
- Función y anatomía del riñónDocumento8 páginasFunción y anatomía del riñónSebastián SeminarioAún no hay calificaciones
- Sistema RenalDocumento102 páginasSistema RenalAdry FaryAún no hay calificaciones
- AnatomíaDocumento5 páginasAnatomíaSanmopus MorpulAún no hay calificaciones
- Histologia Del Sistema Urinario ActualDocumento32 páginasHistologia Del Sistema Urinario ActualAshley MaradiagaAún no hay calificaciones
- Definición Del Sistema UrinarioDocumento8 páginasDefinición Del Sistema UrinarioMichael Joseph EsquivelAún no hay calificaciones
- FISIOLOGIA Del Sistema UrinarioDocumento36 páginasFISIOLOGIA Del Sistema UrinarioMayori Lisseth Ruiz AguirreAún no hay calificaciones
- Sistema RenalDocumento22 páginasSistema RenalcoromotoAún no hay calificaciones
- Anatomía y Fisiología RenalDocumento51 páginasAnatomía y Fisiología RenalMario Nieto'80% (5)
- Sistema Urinario 2021Documento9 páginasSistema Urinario 2021Cristofer Oliver Peñaloza YujraAún no hay calificaciones
- Tarea Del Primer Parcial de GenitourinarioDocumento9 páginasTarea Del Primer Parcial de GenitourinarioRetdiniAún no hay calificaciones
- DiureticosDocumento17 páginasDiureticosFernando Andre AlvarezAún no hay calificaciones
- El Sistema RenalDocumento5 páginasEl Sistema RenaltorukuwhiteAún no hay calificaciones
- Universidad Tecnica Privada Cosmos : El Aparato UrinarioDocumento12 páginasUniversidad Tecnica Privada Cosmos : El Aparato UrinarioMargarita YampiriAún no hay calificaciones
- Tema 5. Sistema Urinario: Excreción UrinariaDocumento7 páginasTema 5. Sistema Urinario: Excreción UrinariaAngela CooperAún no hay calificaciones
- Anatomía y Fisiología Del Aparato GenitourinarioDocumento41 páginasAnatomía y Fisiología Del Aparato Genitourinarioluan navAún no hay calificaciones
- Anatomía y Fisiología RenalDocumento4 páginasAnatomía y Fisiología RenalCarolina DiazAún no hay calificaciones
- Introducción Básica a la Microcirculación de la Sangre: La Siguiente Generación en Cuidados de la SaludDe EverandIntroducción Básica a la Microcirculación de la Sangre: La Siguiente Generación en Cuidados de la SaludAún no hay calificaciones
- Coagulación Para Todos: Medicina Para TodosDe EverandCoagulación Para Todos: Medicina Para TodosCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Banco de PreguntasDocumento43 páginasBanco de PreguntasMilton Siles100% (4)
- Diuretic OsDocumento45 páginasDiuretic OsBelzabeth Carolina Vivas OlivaresAún no hay calificaciones
- Fármacos Que Actual A Nivel Cardiovascular y RenalDocumento34 páginasFármacos Que Actual A Nivel Cardiovascular y RenalsandraAún no hay calificaciones
- Capítulo 3, RenalDocumento56 páginasCapítulo 3, RenalSofía Gutiérrez VictoriaAún no hay calificaciones
- SISTEMA UrinarioDocumento27 páginasSISTEMA UrinarioJuan Carlos DOrso100% (1)
- Fisiologia RenalDocumento15 páginasFisiologia RenalJimena Requejo BustamanteAún no hay calificaciones
- Tema 14. Circulación Renal. Filtración Glomerular.Documento25 páginasTema 14. Circulación Renal. Filtración Glomerular.Pilar del Pino86% (7)
- Semana 3 FISIOLOGIA IVDocumento7 páginasSemana 3 FISIOLOGIA IVArturo Hernandez OlivaAún no hay calificaciones
- Sistema RenalDocumento102 páginasSistema RenalAdry FaryAún no hay calificaciones
- Sistema UrinarioDocumento30 páginasSistema UrinarioEduardo DavilaAún no hay calificaciones
- Líquidos corporales y formación de la orinaDocumento20 páginasLíquidos corporales y formación de la orinaIsaac DerramonaAún no hay calificaciones
- Agentes DiureticosDocumento30 páginasAgentes Diureticossh21005Aún no hay calificaciones
- Taller Sistema UrinarioDocumento5 páginasTaller Sistema UrinarioStephiis CardenasAún no hay calificaciones
- FurosemidaDocumento5 páginasFurosemidaBrian GomezAún no hay calificaciones
- Taller Sistema Excretor Humano 45Documento3 páginasTaller Sistema Excretor Humano 45JuanGuillermoUruetaSierra100% (1)
- Funcion RenalDocumento32 páginasFuncion Renalliz thacharAún no hay calificaciones
- NefronaDocumento5 páginasNefronaDiego RiosAún no hay calificaciones
- Diuréticos del asa de Henle: mecanismo, efectos y usosDocumento3 páginasDiuréticos del asa de Henle: mecanismo, efectos y usosdeyviAún no hay calificaciones
- 2.NEFROLOGIA H.B. DesbloqueadoDocumento144 páginas2.NEFROLOGIA H.B. DesbloqueadoNadith CarballoAún no hay calificaciones
- Farmacos Borrador GoogleDocumento58 páginasFarmacos Borrador GoogleSammy SarangoAún no hay calificaciones
- Comportamiento de La Furosemida en Ratas. FINALDocumento10 páginasComportamiento de La Furosemida en Ratas. FINALJessica GranizoAún no hay calificaciones
- Taller de RenalDocumento8 páginasTaller de RenalRondón Q' VaneAún no hay calificaciones
- Canales Intercambiadores y Bombas de La NefronaDocumento4 páginasCanales Intercambiadores y Bombas de La NefronaJose Fernando Palma RamirezAún no hay calificaciones
- AparatoUrinarioDocumento30 páginasAparatoUrinarioFátima FerreyraAún no hay calificaciones
- Produccon de Orina Diluida y ConcentradaDocumento4 páginasProduccon de Orina Diluida y ConcentradaAna Beatriz Llanos GemioAún no hay calificaciones
- Informe Fisio (Morfo 2) Semana 10Documento14 páginasInforme Fisio (Morfo 2) Semana 10Escoly AltamiranoAún no hay calificaciones
- Histologia Del RiñonDocumento137 páginasHistologia Del RiñonMaa ArliiAún no hay calificaciones
- Seminario Renal II - Sodio y PotasioDocumento65 páginasSeminario Renal II - Sodio y PotasioCayque OliveiraAún no hay calificaciones
- Rev Chil Estud Med V6S1 PDFDocumento119 páginasRev Chil Estud Med V6S1 PDFTomas YungeAún no hay calificaciones
- Taller Digestivo DefinitivoDocumento28 páginasTaller Digestivo DefinitivoEyder Alexander RodriguezAún no hay calificaciones