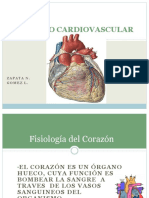
También podría gustarte
- Mecanismos Basicos de La Funcion RenalDocumento3 páginasMecanismos Basicos de La Funcion RenalJuan Jose BarretoAún no hay calificaciones
- Fisiologia Apr Renal 20Documento12 páginasFisiologia Apr Renal 20Victor Hugo Rebaza EsquenAún no hay calificaciones
- Anatomía y Fisiología RenalDocumento51 páginasAnatomía y Fisiología RenalMario Nieto'80% (5)
- Trastornos de La Función GlomerularDocumento133 páginasTrastornos de La Función GlomerularMaster AlonzoAún no hay calificaciones
- Bio 7Documento19 páginasBio 7Benji Pineda CoronelAún no hay calificaciones
- Anatomía y Fisiología RenalDocumento12 páginasAnatomía y Fisiología RenalGrecia HibarraAún no hay calificaciones
- Anatomía y Fisiología de La Función RenalDocumento39 páginasAnatomía y Fisiología de La Función RenalJorge A Uc GuerraAún no hay calificaciones
- Disección Del RiñonDocumento13 páginasDisección Del RiñonDanny Troya GomezAún no hay calificaciones
- Describa La Anatomía Básica Del RiñónDocumento5 páginasDescriba La Anatomía Básica Del RiñónMarcosAntonioAlvaradoAún no hay calificaciones
- Sistema RenalDocumento18 páginasSistema RenalAna LuleAún no hay calificaciones
- Renal IntroduccionDocumento60 páginasRenal IntroduccionINGRID JOHANNA CUESTA JIMENEZAún no hay calificaciones
- 2 - Sistema RenalDocumento43 páginas2 - Sistema RenalBárbara ValdésAún no hay calificaciones
- Anatomia y Fisiologia RenalDocumento26 páginasAnatomia y Fisiologia RenalChristian Montiel100% (1)
- El Aparato UrinarioDocumento8 páginasEl Aparato UrinarionikocjsAún no hay calificaciones
- Anatomia y Fisiologia Renal para EnfermerasDocumento172 páginasAnatomia y Fisiologia Renal para EnfermerasMaria Camila Ramírez Gutiérrez100% (1)
- APARATO URINARIO Resumen Tortora FisiologiaDocumento14 páginasAPARATO URINARIO Resumen Tortora Fisiologiafede12367% (3)
- Fisiologia RenalDocumento19 páginasFisiologia RenalgoodoyvictoriaAún no hay calificaciones
- Taller Sistema Renal de AnatomiaDocumento18 páginasTaller Sistema Renal de AnatomiaJohnatan Vera Moná0% (1)
- Sistema RenalDocumento4 páginasSistema RenalLuis armando Santana tapiaAún no hay calificaciones
- Fisiología Renal y AnatomiaDocumento41 páginasFisiología Renal y AnatomiaAbraxas72Aún no hay calificaciones
- Cápsula Filtrado RenalDocumento8 páginasCápsula Filtrado RenalagustinaAún no hay calificaciones
- Apunte46 Anatomía y Funciones Básicas Del Aparato UrinarioDocumento6 páginasApunte46 Anatomía y Funciones Básicas Del Aparato UrinarioDANIEL ALEJANDROAún no hay calificaciones
- Fisiologia Del Aparato Urinario 1a ClaseDocumento49 páginasFisiologia Del Aparato Urinario 1a ClaseRodrigo S. AvilésAún no hay calificaciones
- Sistema UrinarioDocumento7 páginasSistema Urinarioemil obeidAún no hay calificaciones
- Anatomía y Fisiología RenalDocumento10 páginasAnatomía y Fisiología RenalJoan william Salazar paguasiAún no hay calificaciones
- Sistema ExcretorDocumento38 páginasSistema ExcretorMagui CoralAún no hay calificaciones
- Fisiología Del RiñonDocumento8 páginasFisiología Del RiñonCD MDAún no hay calificaciones
- Aparato UrinarioDocumento64 páginasAparato UrinarioSalvador BARRON MELOAún no hay calificaciones
- Aparato UrinarioDocumento19 páginasAparato UrinarioAnaAún no hay calificaciones
- Excretor - UrinarioDocumento42 páginasExcretor - UrinarioconyypompeiiAún no hay calificaciones
- Aparato Urinario Anatomia y FisiologiaDocumento11 páginasAparato Urinario Anatomia y Fisiologiayordane quispe delgadoAún no hay calificaciones
- Anatomia y Fisiologia RenalDocumento12 páginasAnatomia y Fisiologia RenalCarlos Gonzalez AlemanAún no hay calificaciones
- Sistema RenalDocumento7 páginasSistema RenalHeydi Cardeno PAún no hay calificaciones
- Renal HidrosalinoDocumento37 páginasRenal HidrosalinoViviana Hernandez QuijanoAún no hay calificaciones
- Sistema UrinarioDocumento35 páginasSistema Urinariohenriquezrodriguez2023Aún no hay calificaciones
- M1 T1 Actualización Sobre Enfermedad Renal PDFDocumento10 páginasM1 T1 Actualización Sobre Enfermedad Renal PDFFernando SantaellaAún no hay calificaciones
- El Aparato UrinarioDocumento42 páginasEl Aparato Urinariovaleria isabel poot haazAún no hay calificaciones
- Clase 1 y 2Documento43 páginasClase 1 y 2studyshop29Aún no hay calificaciones
- Sistema Urinario - BrendaDocumento40 páginasSistema Urinario - BrendaBrenda TrujilloAún no hay calificaciones
- Sistema Urinario OficialDocumento87 páginasSistema Urinario OficialĶéýáñ Jén ĶínsAún no hay calificaciones
- Folleto Complementario de NefrologíaDocumento14 páginasFolleto Complementario de NefrologíaOffman VacaAún no hay calificaciones
- RNALDocumento16 páginasRNALEVELYN GONZALESAún no hay calificaciones
- Monografia Sistema UrinarioDocumento14 páginasMonografia Sistema UrinarioAlexander Herrera67% (6)
- Renal - Tarea 1-5Documento31 páginasRenal - Tarea 1-5barrioslissette87Aún no hay calificaciones
- Resumen Bloque ViDocumento7 páginasResumen Bloque ViAlba WAún no hay calificaciones
- CLASE No 14 Fisiologia RenalDocumento26 páginasCLASE No 14 Fisiologia RenalAnderson BenitesAún no hay calificaciones
- RenalDocumento32 páginasRenalMacarena RamírezAún no hay calificaciones
- Guia Seminario Dialisis y HemodialisisDocumento15 páginasGuia Seminario Dialisis y Hemodialisisjazmin echeverria peñarandaAún no hay calificaciones
- Fisiología RenalDocumento32 páginasFisiología RenalcarmenAún no hay calificaciones
- Guía Nº2. Fisiología RenalDocumento17 páginasGuía Nº2. Fisiología RenalBorreguinho OrregoAún no hay calificaciones
- Sistema UrinarioDocumento32 páginasSistema Urinariofernanda tinajero100% (1)
- NefrologíaDocumento59 páginasNefrologíakarlasifontesAún no hay calificaciones
- Histologia Del Sistema Urinario ActualDocumento32 páginasHistologia Del Sistema Urinario ActualAshley MaradiagaAún no hay calificaciones
- Taller Semio RenalDocumento7 páginasTaller Semio RenalWendy Victoria Castillo BenitezAún no hay calificaciones
- Anatomia de Los RiñonesDocumento6 páginasAnatomia de Los RiñonesAlexandre QuisbertAún no hay calificaciones
- Anatomía y Fisiología Del Sistema UrinarioDocumento7 páginasAnatomía y Fisiología Del Sistema UrinarioMarcos LeonAún no hay calificaciones
- Introducción Básica a la Microcirculación de la Sangre: La Siguiente Generación en Cuidados de la SaludDe EverandIntroducción Básica a la Microcirculación de la Sangre: La Siguiente Generación en Cuidados de la SaludAún no hay calificaciones
- Coagulación Para Todos: Medicina Para TodosDe EverandCoagulación Para Todos: Medicina Para TodosCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Piensa Antes De Beber: Las consecuencias de consumir bebidas alcohólicasDe EverandPiensa Antes De Beber: Las consecuencias de consumir bebidas alcohólicasCalificación: 5 de 5 estrellas5/5 (1)
- 4.hemoglobina TeoríaDocumento17 páginas4.hemoglobina TeoríaJasna ZovakAún no hay calificaciones
- Organización FuncionalDocumento4 páginasOrganización FuncionalJasna ZovakAún no hay calificaciones
- CardiovascularDocumento11 páginasCardiovascularJasna ZovakAún no hay calificaciones
- Sist. Endócrino y Sus TrastornosDocumento2 páginasSist. Endócrino y Sus TrastornosJasna ZovakAún no hay calificaciones
- 12 Fisiología Sist. NerviosoDocumento10 páginas12 Fisiología Sist. NerviosoJasna ZovakAún no hay calificaciones
- SNC - Cuadros ResumenDocumento5 páginasSNC - Cuadros ResumenJasna ZovakAún no hay calificaciones
- LeucemiasDocumento39 páginasLeucemiasJasna ZovakAún no hay calificaciones
- Proceso ReproductivoDocumento8 páginasProceso ReproductivoJasna ZovakAún no hay calificaciones
- Sistema Nervioso y Org. de Los SentidosDocumento63 páginasSistema Nervioso y Org. de Los SentidosJasna ZovakAún no hay calificaciones
- 10 Fisio Del Ap. Rep. MasculinoDocumento2 páginas10 Fisio Del Ap. Rep. MasculinoJasna ZovakAún no hay calificaciones
- 8 Introducción A MetabolismoDocumento18 páginas8 Introducción A MetabolismoJasna ZovakAún no hay calificaciones
- Sistema Muscular Intro 13Documento3 páginasSistema Muscular Intro 13Jasna ZovakAún no hay calificaciones
- 7 Sistema Excretor - ResumenDocumento5 páginas7 Sistema Excretor - ResumenJasna ZovakAún no hay calificaciones
- 3 Fisiología SangreDocumento28 páginas3 Fisiología SangreJasna ZovakAún no hay calificaciones
- 9 Fisiología AP. ReproductorDocumento6 páginas9 Fisiología AP. ReproductorJasna ZovakAún no hay calificaciones
- Sistema Linfático Intro 13Documento2 páginasSistema Linfático Intro 13Jasna ZovakAún no hay calificaciones
- SangreDocumento8 páginasSangreJasna ZovakAún no hay calificaciones
- Sistema Articular Intro 13Documento2 páginasSistema Articular Intro 13Jasna ZovakAún no hay calificaciones
- Guía Célula y FuncionesDocumento2 páginasGuía Célula y FuncionesJasna ZovakAún no hay calificaciones
- ESQUELETO Intro 13Documento3 páginasESQUELETO Intro 13Jasna ZovakAún no hay calificaciones
- Sistema Circulatorio Intro 13Documento4 páginasSistema Circulatorio Intro 13Jasna ZovakAún no hay calificaciones
- Guía Esqueleto HumanoDocumento2 páginasGuía Esqueleto HumanoJasna ZovakAún no hay calificaciones
- 11.citometria de FlujoDocumento1 página11.citometria de FlujoJasna ZovakAún no hay calificaciones
- GlosarioDocumento2 páginasGlosarioJasna ZovakAún no hay calificaciones
- 10.leucemias TeoriaDocumento13 páginas10.leucemias TeoriaJasna ZovakAún no hay calificaciones
- 3.el Hierro Es Un Elemento Esencial para La VidaDocumento7 páginas3.el Hierro Es Un Elemento Esencial para La VidaJasna ZovakAún no hay calificaciones
- 6 HematopoyesisDocumento45 páginas6 HematopoyesisJasna ZovakAún no hay calificaciones
- 9.tipos de AnemiasDocumento23 páginas9.tipos de AnemiasJasna ZovakAún no hay calificaciones
- 2 HemogramaDocumento23 páginas2 HemogramaJasna ZovakAún no hay calificaciones
- Sintesis de ProteinasDocumento9 páginasSintesis de ProteinasHECTOR AVENDAÑO RAMOSAún no hay calificaciones
- Conferencia Citoplasma Completo OkDocumento60 páginasConferencia Citoplasma Completo OkPawan Alvarez PerezAún no hay calificaciones
- Gymkana Nervioso 2 - 20231128 - 112203 - 0000Documento5 páginasGymkana Nervioso 2 - 20231128 - 112203 - 0000Matias ValenzuelaAún no hay calificaciones
- Guia de TP 9°Documento13 páginasGuia de TP 9°Ser leAún no hay calificaciones
- Aparato Digestivo y PielDocumento9 páginasAparato Digestivo y Pielmichelle ordoñezAún no hay calificaciones
- Valorización de La Naturaleza y El Territorio. Opciones Teóricas-Metodológicas para Pensar Otras Territorialidades PosiblesDocumento16 páginasValorización de La Naturaleza y El Territorio. Opciones Teóricas-Metodológicas para Pensar Otras Territorialidades PosiblesLore CoppiaroloAún no hay calificaciones
- Organizador de MurrayDocumento1 páginaOrganizador de MurrayKenia AguilarAún no hay calificaciones
- El Puesto Del Hombre en La NaturalezaDocumento2 páginasEl Puesto Del Hombre en La NaturalezaZarate Mejía AlexisAún no hay calificaciones
- Problemas Genetica 2022Documento2 páginasProblemas Genetica 2022Gloria Elena RojasAún no hay calificaciones
- El Líder Resonante Crea Más - Resumen Pers.Documento4 páginasEl Líder Resonante Crea Más - Resumen Pers.SILVIA TORRESAún no hay calificaciones
- Biologia Del DesarrolloDocumento18 páginasBiologia Del DesarrollocristianAún no hay calificaciones
- Adicionales - 6°Documento53 páginasAdicionales - 6°CUSSIAún no hay calificaciones
- Agar CelulosaDocumento2 páginasAgar CelulosaEstefania Mendez100% (1)
- 14 Práctica Histología - Cavidad Bucal 2 2022 - 1Documento30 páginas14 Práctica Histología - Cavidad Bucal 2 2022 - 1sofia GAún no hay calificaciones
- UD 2. Tema 3. División EuglenophytaDocumento32 páginasUD 2. Tema 3. División EuglenophytairulagunAún no hay calificaciones
- Método EtnográficoDocumento31 páginasMétodo EtnográficoMaria Dolores Torres MendozaAún no hay calificaciones
- Evaluación Ciencias Unidad 2 Los Seres Vivos. Primeros.Documento6 páginasEvaluación Ciencias Unidad 2 Los Seres Vivos. Primeros.Gisela MartinezAún no hay calificaciones
- Biometria Pez NativoDocumento7 páginasBiometria Pez NativoJosé CPAún no hay calificaciones
- Guia Culivo Yuca 1Documento1 páginaGuia Culivo Yuca 1jesus carlosamaAún no hay calificaciones
- Proyecto 2Documento8 páginasProyecto 2almaAún no hay calificaciones
- Una Nueva Especie de Leucheria (AsteraceaeDocumento6 páginasUna Nueva Especie de Leucheria (AsteraceaeAlejandro RiquelmeAún no hay calificaciones
- CCNN 8vo Unidad 2Documento5 páginasCCNN 8vo Unidad 2Nieves Maria Maria Santarrosa CarguaAún no hay calificaciones
- Clasificados 04.02Documento4 páginasClasificados 04.02Nicolás Dante Di FioriAún no hay calificaciones
- UNDP PY 2020 Componente Biodiversidad ItapuaDocumento146 páginasUNDP PY 2020 Componente Biodiversidad ItapuaGilceAún no hay calificaciones
- El OjoDocumento9 páginasEl OjoEstephany SantosAún no hay calificaciones
- Nominas de 4to Año 2020-2021Documento17 páginasNominas de 4to Año 2020-2021Salvador MartinezAún no hay calificaciones
- Entrevista Kin SicaDocumento113 páginasEntrevista Kin SicaMiguelAún no hay calificaciones
- Informe de InvertebradosDocumento8 páginasInforme de InvertebradosJose CantareroAún no hay calificaciones
- 2.tema: Interacciones: S E C U N D A R I ADocumento6 páginas2.tema: Interacciones: S E C U N D A R I AJandi RosetAún no hay calificaciones
- Histofisiologia PeriapicalDocumento14 páginasHistofisiologia PeriapicalLia YepezAún no hay calificaciones