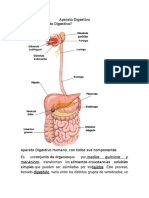
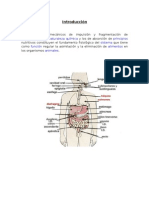
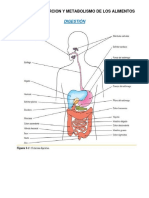
También podría gustarte
- Guía básica de alimentación y nutriciónDe EverandGuía básica de alimentación y nutriciónCalificación: 5 de 5 estrellas5/5 (2)
- Anatomía y Fiisologia Del Aparato DigestivoDocumento12 páginasAnatomía y Fiisologia Del Aparato DigestivoYahir IzquierdoAún no hay calificaciones
- Felampe 1 - MergedDocumento8 páginasFelampe 1 - MergedSergio Retamal MirandaAún no hay calificaciones
- DigestiónDocumento10 páginasDigestiónAlan PonceAún no hay calificaciones
- Taller Bioquimica DigestionDocumento8 páginasTaller Bioquimica DigestionESTEBAN BONILLA CHINCHILLAAún no hay calificaciones
- Digestion Absorcion MetabolismoDocumento12 páginasDigestion Absorcion MetabolismojeniferAún no hay calificaciones
- Digestión - Wikipedia, La Enciclopedia LibreDocumento66 páginasDigestión - Wikipedia, La Enciclopedia LibreCarlosAún no hay calificaciones
- 1er Parcial - Resumenes de Nutrición - Dra. Emen.Documento54 páginas1er Parcial - Resumenes de Nutrición - Dra. Emen.Adriana DelgadoAún no hay calificaciones
- Aparato DigestivoDocumento12 páginasAparato DigestivoIzselaDleonAún no hay calificaciones
- Bibliografía - UNIDAD 3 - ANATOMIADocumento10 páginasBibliografía - UNIDAD 3 - ANATOMIAcarolinaAún no hay calificaciones
- CEF Bases Biologicas y Fisiologicas Del Entrenamiento Clase 3 PDFDocumento19 páginasCEF Bases Biologicas y Fisiologicas Del Entrenamiento Clase 3 PDFanamoreno8Aún no hay calificaciones
- Fisiopatologia de Intestino DelgadoDocumento59 páginasFisiopatologia de Intestino DelgadoROSSYAún no hay calificaciones
- Resumen SISTEMA - DIGESTIVO - PDFDocumento26 páginasResumen SISTEMA - DIGESTIVO - PDFLina Cordero BerrioAún no hay calificaciones
- Biofisica +Documento14 páginasBiofisica +DayaAún no hay calificaciones
- Informe Biologia.. S. Digestivo, Circulatorio, RespiratorioDocumento29 páginasInforme Biologia.. S. Digestivo, Circulatorio, RespiratorioMaickol F BalarezoAún no hay calificaciones
- Digestion y Absorcion 22Documento18 páginasDigestion y Absorcion 22Fiorela NavarroAún no hay calificaciones
- Sistema DigestivoDocumento11 páginasSistema DigestivoAmericanAún no hay calificaciones
- GUIA DigestiónDocumento9 páginasGUIA Digestiónjanny RomeroAún no hay calificaciones
- Sistema Gastrointestinal (SGI)Documento23 páginasSistema Gastrointestinal (SGI)VanessaLevanoPonteAún no hay calificaciones
- TRABAJO DE ANATOMIA - CuestionarioDocumento57 páginasTRABAJO DE ANATOMIA - CuestionarionathyvicAún no hay calificaciones
- DigestiónDocumento5 páginasDigestiónluciaAún no hay calificaciones
- Biosfera DigestivoDocumento10 páginasBiosfera DigestivoAngeles ArnoAún no hay calificaciones
- Digestión y Absorción: Definiciones y Conceptos ClavesDocumento28 páginasDigestión y Absorción: Definiciones y Conceptos ClavesBrayan 34Aún no hay calificaciones
- Absorción y Digestión de Macronutrientes en El Intestino DelgadoDocumento9 páginasAbsorción y Digestión de Macronutrientes en El Intestino DelgadoNATALIA ROXANA CONCILCO JUAREZAún no hay calificaciones
- Aparato DigestivoDocumento15 páginasAparato Digestivogiselle BasualdoAún no hay calificaciones
- Lleno Actividad 4 - Proceso Digestivo HumanoDocumento7 páginasLleno Actividad 4 - Proceso Digestivo Humanoricardo garciaAún no hay calificaciones
- Clase 2Documento5 páginasClase 2Emer Yvan Horna CadenillasAún no hay calificaciones
- Informe de FisicoquimicaDocumento5 páginasInforme de FisicoquimicaBetzy Valery Poma PerezAún no hay calificaciones
- Cuestionario de Evaluación de La Función Gastrointestinal BuenoDocumento16 páginasCuestionario de Evaluación de La Función Gastrointestinal BuenoLirio LópezAún no hay calificaciones
- Guia - de - Estudio - de - La - Unidad Digestion y Absorciã - N de Nutrientes y de Bioenergetica 2020.1Documento33 páginasGuia - de - Estudio - de - La - Unidad Digestion y Absorciã - N de Nutrientes y de Bioenergetica 2020.1Edersson ReyesAún no hay calificaciones
- DigestivoDocumento7 páginasDigestivoDaiana ValdiviesoAún no hay calificaciones
- Sistema Digestivo HumanoDocumento8 páginasSistema Digestivo HumanoJOSE CORREAAún no hay calificaciones
- Fisiología Del Aparato Digestivo: ¿Qué Es La Digestión?Documento18 páginasFisiología Del Aparato Digestivo: ¿Qué Es La Digestión?Jhonatan Quispe HuancaAún no hay calificaciones
- Compilacion Desarrollo Nutricional de La ComunidadDocumento25 páginasCompilacion Desarrollo Nutricional de La ComunidadJocelyn RamirezAún no hay calificaciones
- Biologia 1 ObjetivoDocumento10 páginasBiologia 1 Objetivovelasquezgilbelys547Aún no hay calificaciones
- Foro Semana 13 - Fisiología DigestivaDocumento6 páginasForo Semana 13 - Fisiología DigestivaYomira YulissaAún no hay calificaciones
- Compilacion Desarrollo Nutricional de La ComunidadDocumento31 páginasCompilacion Desarrollo Nutricional de La ComunidadAldo TorresAún no hay calificaciones
- DigestionDocumento15 páginasDigestionfrank1605Aún no hay calificaciones
- Sistema DigestivoDocumento7 páginasSistema DigestivoJose A. Beltrán BolañosAún no hay calificaciones
- Introducción Al Sistema DigestivoDocumento47 páginasIntroducción Al Sistema DigestivoLuz Alejandra Ruelas YanesAún no hay calificaciones
- Digestivo Vane-AnaDocumento18 páginasDigestivo Vane-AnaDaiana tabordaAún no hay calificaciones
- Resumen BromatologiaDocumento9 páginasResumen BromatologiaAnyhuAún no hay calificaciones
- Sistema Digestivo FisiologiaDocumento10 páginasSistema Digestivo FisiologiaJoan william Salazar paguasiAún no hay calificaciones
- Fisiologia Del Sistema Digestivo (Alimentacion y Nutricion) Hoy 13 de NovDocumento131 páginasFisiologia Del Sistema Digestivo (Alimentacion y Nutricion) Hoy 13 de Novjesus perezAún no hay calificaciones
- Clase 2Documento5 páginasClase 2martin lunaAún no hay calificaciones
- Sistema DigestivoDocumento2 páginasSistema DigestivoFernando Aguilar100% (1)
- DigestionDocumento9 páginasDigestionFRANCIA IVETTE ALVAREZ CALDERONAún no hay calificaciones
- Disquinesia BiliarDocumento5 páginasDisquinesia Biliaranon_716919997Aún no hay calificaciones
- Tema 4 Nutrición IDocumento7 páginasTema 4 Nutrición IAMANDA VILLARROEL RAMOSAún no hay calificaciones
- La Nutrición en Los Seres VivosDocumento14 páginasLa Nutrición en Los Seres Vivosilias fakhriAún no hay calificaciones
- Absorción Por Mucosa GastroDocumento5 páginasAbsorción Por Mucosa GastroHector ValdiviaAún no hay calificaciones
- 7mo Brom y Nut Act 2 DigestionDocumento9 páginas7mo Brom y Nut Act 2 Digestionpehuen323Aún no hay calificaciones
- INGESTIÓNDocumento5 páginasINGESTIÓNminipabloxd1Aún no hay calificaciones
- Digestion-Absorcion-MetabolismoDocumento8 páginasDigestion-Absorcion-MetabolismoJeriel Sosa VerasAún no hay calificaciones
- Resumen DigestivoDocumento13 páginasResumen DigestivojosefinaAún no hay calificaciones
- Fisiología Del Aparato DigestivoDocumento5 páginasFisiología Del Aparato DigestivoCitlalli PonceAún no hay calificaciones
- DIGESTIÓNDocumento12 páginasDIGESTIÓNrodofoxAún no hay calificaciones
- Digestion GastricaDocumento19 páginasDigestion GastricaPedro José Barboza EspejoAún no hay calificaciones
- 01-Sistema Digestivo HumanoDocumento42 páginas01-Sistema Digestivo Humanoفاطمة كوريلا100% (1)
- Resumen UD 1,2Documento2 páginasResumen UD 1,2Marcos Belmar OrtizAún no hay calificaciones
- Tema 6. Valoración de Compartimentos CorporalesDocumento37 páginasTema 6. Valoración de Compartimentos CorporalesMarcos Belmar OrtizAún no hay calificaciones
- Actividades UT2 DesarrolloDocumento1 páginaActividades UT2 DesarrolloMarcos Belmar OrtizAún no hay calificaciones
- Tema 9. Alimentación Embarazo y LactanciaDocumento16 páginasTema 9. Alimentación Embarazo y LactanciaMarcos Belmar OrtizAún no hay calificaciones
- Tema 7. Valoración de La Ingesta y Cálculo Balance EnergéticoDocumento13 páginasTema 7. Valoración de La Ingesta y Cálculo Balance EnergéticoMarcos Belmar OrtizAún no hay calificaciones
- Acidez en Leches (Entera y Desnatada)Documento2 páginasAcidez en Leches (Entera y Desnatada)Marcos Belmar OrtizAún no hay calificaciones
- Acidez FijaDocumento2 páginasAcidez FijaMarcos Belmar OrtizAún no hay calificaciones
- CalculosdietaDocumento2 páginasCalculosdietaMarcos Belmar OrtizAún no hay calificaciones
- Acidez Total VinagresDocumento2 páginasAcidez Total VinagresMarcos Belmar OrtizAún no hay calificaciones
- Tema 5. Digestión, Absorción y MetabolismoDocumento36 páginasTema 5. Digestión, Absorción y MetabolismoMarcos Belmar OrtizAún no hay calificaciones
- Acidez Pastas AlimenticiasDocumento2 páginasAcidez Pastas AlimenticiasMarcos Belmar Ortiz100% (1)
- Acidez de Una MermeladaDocumento2 páginasAcidez de Una MermeladaMarcos Belmar OrtizAún no hay calificaciones
- Tema.2 DisolucionesDocumento5 páginasTema.2 DisolucionesMarcos Belmar OrtizAún no hay calificaciones
- FISIOPATOLOGIA5Documento3 páginasFISIOPATOLOGIA5Marcos Belmar OrtizAún no hay calificaciones
- Cálculos Práctica 1Documento8 páginasCálculos Práctica 1Marcos Belmar OrtizAún no hay calificaciones
- Guión Práctica 1Documento20 páginasGuión Práctica 1Marcos Belmar OrtizAún no hay calificaciones
- Deberes Recetas AeqDocumento3 páginasDeberes Recetas AeqMarcos Belmar OrtizAún no hay calificaciones
- Tema 7. Valoración de La Ingesta y Cálculo Balance EnergéticoDocumento13 páginasTema 7. Valoración de La Ingesta y Cálculo Balance EnergéticoMarcos Belmar OrtizAún no hay calificaciones
- Tema 8. Dietas en Adultos SanosDocumento23 páginasTema 8. Dietas en Adultos SanosMarcos Belmar OrtizAún no hay calificaciones
- Tema 6. Valoración de Compartimentos CorporalesDocumento37 páginasTema 6. Valoración de Compartimentos CorporalesMarcos Belmar OrtizAún no hay calificaciones
- TrabajofolDocumento14 páginasTrabajofolMarcos Belmar OrtizAún no hay calificaciones
- Fol 1ºdietética AlumnosDocumento16 páginasFol 1ºdietética AlumnosMarcos Belmar OrtizAún no hay calificaciones
- Tema 3 - Nuevo 2022-23Documento25 páginasTema 3 - Nuevo 2022-23Marcos Belmar OrtizAún no hay calificaciones
- Tema 1,2,3 Matemáticas Calculo DiferencialDocumento78 páginasTema 1,2,3 Matemáticas Calculo DiferencialMarcos Belmar OrtizAún no hay calificaciones
- Preguntas Tipo Test Fisiopatología, SantiagoDocumento2 páginasPreguntas Tipo Test Fisiopatología, SantiagoMarcos Belmar OrtizAún no hay calificaciones
- Vitamina B3Documento5 páginasVitamina B3Marcos Belmar OrtizAún no hay calificaciones
- TEMA2AEQDocumento22 páginasTEMA2AEQMarcos Belmar OrtizAún no hay calificaciones
- TEMA 21 Anabolismo Hetero y AutoDocumento1 páginaTEMA 21 Anabolismo Hetero y AutoMarcos Belmar OrtizAún no hay calificaciones
- T1 - La Imaginación Sociologica SociologíaDocumento62 páginasT1 - La Imaginación Sociologica SociologíaMarcos Belmar OrtizAún no hay calificaciones
- TRABAJOAEQDocumento8 páginasTRABAJOAEQMarcos Belmar OrtizAún no hay calificaciones
- Patologías Del EstomagoDocumento27 páginasPatologías Del EstomagotamarAún no hay calificaciones
- 10 Los MoluscosDocumento141 páginas10 Los MoluscosTeddy Dehesa100% (2)
- Control Nervioso de La Función Gastrointestinal Sistema Nervioso EntéricoDocumento7 páginasControl Nervioso de La Función Gastrointestinal Sistema Nervioso EntéricoOscar Larios AyalaAún no hay calificaciones
- Embriologia en SistemasDocumento12 páginasEmbriologia en SistemasLuis Martinez NavarroAún no hay calificaciones
- 2191 Presentación Electrónica Educativa 2164 1-10-20190405Documento55 páginas2191 Presentación Electrónica Educativa 2164 1-10-20190405GladysAún no hay calificaciones
- EQUINOSDocumento6 páginasEQUINOSChristian Alejandro100% (1)
- Digestivo y ExcretorDocumento59 páginasDigestivo y ExcretorHola soy Kham y no soy Terrorista e.eAún no hay calificaciones
- La Acidez EstomacalDocumento1 páginaLa Acidez EstomacalHerbert Isaac Arce IngaAún no hay calificaciones
- Anatomía e Histología de Faringe, AmígdalasDocumento8 páginasAnatomía e Histología de Faringe, AmígdalasMishelle CastroAún no hay calificaciones
- Medios de Contraste en RadiologiaDocumento16 páginasMedios de Contraste en RadiologiaRobert MoranAún no hay calificaciones
- Tratamiento Quirúrgico Del Reflujo Gastroesofágico Del Adulto Por LaparotomíaDocumento20 páginasTratamiento Quirúrgico Del Reflujo Gastroesofágico Del Adulto Por LaparotomíaDionisio José Ozuna LoaizaAún no hay calificaciones
- Sesion 10 Practica Sistema Digestivo y EstrectorDocumento3 páginasSesion 10 Practica Sistema Digestivo y EstrectorNICOL ESTEFFANI VALVERDE MAZAAún no hay calificaciones
- Informe 1 I - A - CDocumento28 páginasInforme 1 I - A - CLiliana Tello PortillaAún no hay calificaciones
- Nutricion El Mantenimiento de La VidaDocumento10 páginasNutricion El Mantenimiento de La VidaWilson OspinoAún no hay calificaciones
- AyunoDocumento13 páginasAyunoPavel LunaAún no hay calificaciones
- Primera Evaluación Básicas - Dic 13-21Documento21 páginasPrimera Evaluación Básicas - Dic 13-21juanAún no hay calificaciones
- Linfáticos EsplacnologíaDocumento5 páginasLinfáticos EsplacnologíaRamiro EspositoAún no hay calificaciones
- Aparato DigestivoDocumento57 páginasAparato DigestivoKelly FloresAún no hay calificaciones
- ANATOMÍADocumento58 páginasANATOMÍAFatima LopezAún no hay calificaciones
- FrutasDocumento7 páginasFrutasGiovanaAún no hay calificaciones
- 33 Aspirador Gastrico Medica-DDocumento13 páginas33 Aspirador Gastrico Medica-Decko_golden3812Aún no hay calificaciones
- ECOGRAFIADocumento6 páginasECOGRAFIAKtrym BustosAún no hay calificaciones
- Información - GastroparesiaDocumento7 páginasInformación - GastroparesiaPablo G. BledtAún no hay calificaciones
- Fisiología FinalDocumento65 páginasFisiología FinalCamila PiraneoAún no hay calificaciones
- Sistema GastrointestinalDocumento21 páginasSistema GastrointestinalMaría C MoralesAún no hay calificaciones
- Parciales Cirugía 2022Documento43 páginasParciales Cirugía 2022APANQUI QUILLA VANEZAAún no hay calificaciones
- Apunte Sistema Digestivo HistologiaDocumento12 páginasApunte Sistema Digestivo HistologiaseahhAún no hay calificaciones
- 1 Cuaderno Purpura EnergiasDocumento13 páginas1 Cuaderno Purpura EnergiasAlanna BrownAún no hay calificaciones
- Imprimir Guia 2 Ciencias 8ºDocumento4 páginasImprimir Guia 2 Ciencias 8ºJadore34Aún no hay calificaciones
- Examen Internos CirugiaDocumento13 páginasExamen Internos CirugiaPaula LoretoAún no hay calificaciones