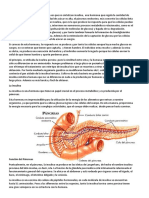
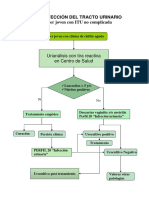
También podría gustarte
- Hemoglobin A 3Documento32 páginasHemoglobin A 3Mirelli Duran100% (1)
- Albumina 2 1Documento2 páginasAlbumina 2 1Yadira Hernandez Ramirez100% (1)
- Hemoglobina PDFDocumento9 páginasHemoglobina PDFJavi R. HatemAún no hay calificaciones
- 1 Unidad Feto PlacentariaDocumento42 páginas1 Unidad Feto Placentariakatty velaAún no hay calificaciones
- Metabolismo de La HBDocumento16 páginasMetabolismo de La HBRosario LagunaAún no hay calificaciones
- La Secreción de La InsulinaDocumento5 páginasLa Secreción de La Insulinaeloydc17Aún no hay calificaciones
- El Riñón y GlucosaDocumento16 páginasEl Riñón y Glucosavicgzz100% (1)
- Psicología CristianaDocumento5 páginasPsicología CristianaErikkson GuzmanAún no hay calificaciones
- Hemoglobina FetalDocumento3 páginasHemoglobina FetalPayola TitoAún no hay calificaciones
- Acidosis Metabolica en UciDocumento34 páginasAcidosis Metabolica en UciArturo1985A100% (1)
- Acido Fólico - SeminarioDocumento26 páginasAcido Fólico - SeminarioohgalemanAún no hay calificaciones
- Metabolismo Del CalcioDocumento41 páginasMetabolismo Del CalcioionatnllAún no hay calificaciones
- Enfermedad de WolmanDocumento4 páginasEnfermedad de WolmanNerylyn GonzálezAún no hay calificaciones
- Bioquimica Proteinas Plasmaticas TerminadoDocumento15 páginasBioquimica Proteinas Plasmaticas Terminadoerick marioAún no hay calificaciones
- Bioquimica MédicaDocumento2 páginasBioquimica MédicaVictor Arce Colque100% (1)
- Caso Clinico GastroenterologiaDocumento23 páginasCaso Clinico GastroenterologiaAlexandra Pauccar Rendon100% (1)
- Nutricion en El EmbarazoDocumento15 páginasNutricion en El EmbarazoAlison TapiaAún no hay calificaciones
- Metabolismo de Glucosa Fructusa y GalactosaDocumento27 páginasMetabolismo de Glucosa Fructusa y GalactosaLorena DuránAún no hay calificaciones
- Deficit de Piruvato CarboxilsaDocumento3 páginasDeficit de Piruvato CarboxilsaAndrea Stefanía CuevaAún no hay calificaciones
- Insulina GlucagonDocumento24 páginasInsulina GlucagonAugusto Polo Casale Lara0% (1)
- Lipo Protein AsDocumento7 páginasLipo Protein Aseliana bustamante100% (1)
- Caso Clinico 1 (ERC, DM2 y HTA)Documento19 páginasCaso Clinico 1 (ERC, DM2 y HTA)Jose Roman Mauricio PanduroAún no hay calificaciones
- Anemia DrepanocíticaDocumento3 páginasAnemia DrepanocíticaJhon MontañoAún no hay calificaciones
- Hemoglobina Curva de DisociacionDocumento55 páginasHemoglobina Curva de DisociacionDaniel Farfan Pujadas100% (2)
- Expo - Diabetes Mellitus PDFDocumento184 páginasExpo - Diabetes Mellitus PDFCristhian SandovalAún no hay calificaciones
- Fisiopatologia DiabetesDocumento9 páginasFisiopatologia DiabetesJUDITH CAAMAÑOAún no hay calificaciones
- Porfiri AsDocumento31 páginasPorfiri Asnatalia100% (1)
- Complicaciones PostransfusionalesDocumento25 páginasComplicaciones Postransfusionalesww.pacourgencias.blogspot.comAún no hay calificaciones
- Anemia MegaloblasticaDocumento40 páginasAnemia MegaloblasticaNayaritaOtianaPalominoMiravalAún no hay calificaciones
- De Donde Proviene La InsulinaDocumento3 páginasDe Donde Proviene La InsulinaIngeniero GregoryAún no hay calificaciones
- Campylobacter JejuniDocumento18 páginasCampylobacter Jejuniabc abcAún no hay calificaciones
- Vitamina K 2021Documento63 páginasVitamina K 2021Endre CordovaAún no hay calificaciones
- Ferro, Hemoglobina e Bilirrubina - 23472 V Octubre 2013 (1) - 2Documento64 páginasFerro, Hemoglobina e Bilirrubina - 23472 V Octubre 2013 (1) - 2Karen SánchezAún no hay calificaciones
- Hemoglobina BajaDocumento2 páginasHemoglobina Bajaomatus99Aún no hay calificaciones
- Enfermedad GaucherDocumento27 páginasEnfermedad GaucherAlex GualliAún no hay calificaciones
- Carbohidratos ConejoDocumento5 páginasCarbohidratos Conejomonica100% (1)
- GlomerulonefritisDocumento3 páginasGlomerulonefritisDeyanira MezaAún no hay calificaciones
- GluconeogenesisDocumento20 páginasGluconeogenesisdan100% (2)
- Vitamina Complejo BDocumento5 páginasVitamina Complejo Banon_377570667Aún no hay calificaciones
- Rutas Metabólicas - Leones - Por - La - SaludDocumento25 páginasRutas Metabólicas - Leones - Por - La - SaludALAN FERNANDO GONZALEZ SERRANOAún no hay calificaciones
- Principales Síndromes Del Sistema DigestivoDocumento8 páginasPrincipales Síndromes Del Sistema DigestivoAnny Andreina Rengel RodriguezAún no hay calificaciones
- Cuadro Comparativo de Enfermedades LisosomalesDocumento4 páginasCuadro Comparativo de Enfermedades Lisosomaleseduardo quirozAún no hay calificaciones
- Fisiopatologia Higado 202010Documento51 páginasFisiopatologia Higado 202010DariforiAún no hay calificaciones
- Enfermedad de KrabbeDocumento15 páginasEnfermedad de KrabbeRicardo MembreñoAún no hay calificaciones
- Anemia FerropenicaDocumento13 páginasAnemia FerropenicaMaria Mendoza100% (1)
- Determinacion TrigliceridosDocumento3 páginasDeterminacion TrigliceridosCarlos Jesus Lachira ZamoraAún no hay calificaciones
- Beriberi .Documento23 páginasBeriberi .Renzo Terroba Yzaguirre100% (1)
- Embriología 8Documento51 páginasEmbriología 8Nicole DíazAún no hay calificaciones
- Dislipidemia Mixta Severa en PoblaciónDocumento12 páginasDislipidemia Mixta Severa en Poblaciónfadyth arizaAún no hay calificaciones
- SEMANA 8 - Función Hepática BioquímicaDocumento8 páginasSEMANA 8 - Función Hepática BioquímicaJuan David Mosos CamposAún no hay calificaciones
- Encefalopatía HepáticaDocumento51 páginasEncefalopatía HepáticaPaulichie100% (11)
- Indicadores Biométricos y Bioquímicos en La Enfermedad Por Hígado Graso No AlcohólicoDocumento17 páginasIndicadores Biométricos y Bioquímicos en La Enfermedad Por Hígado Graso No AlcohólicoOdalys RicardoAún no hay calificaciones
- Cuestionario Sobre Proteinas y EnzimasDocumento8 páginasCuestionario Sobre Proteinas y EnzimasLady bloggersAún no hay calificaciones
- Esquema de Alimentación en Mujeres EmbarazadasDocumento1 páginaEsquema de Alimentación en Mujeres EmbarazadasColandLechugita MixAún no hay calificaciones
- Ensayo GlucolisisDocumento10 páginasEnsayo GlucolisisPame Salinas BuenañoAún no hay calificaciones
- Caso Clinico 1 Anemia MegaloblasticaDocumento25 páginasCaso Clinico 1 Anemia MegaloblasticahaferAún no hay calificaciones
- S11 Laboratorio BioquimicaDocumento14 páginasS11 Laboratorio BioquimicaKia ShocushAún no hay calificaciones
- Anemias HemoliticasDocumento14 páginasAnemias HemoliticasSandra Gabriela Guzman GallegosAún no hay calificaciones
- Metabolismo Del Acido Folico y Vitamina b12Documento5 páginasMetabolismo Del Acido Folico y Vitamina b12Felipe A. Besoain CarocaAún no hay calificaciones
- Fisiología de La HemoglobinaDocumento10 páginasFisiología de La HemoglobinaHenryk BazalarAún no hay calificaciones
- Programacion de Programas Preventivos PromocionalesDocumento30 páginasProgramacion de Programas Preventivos Promocionalesapi-287812609100% (1)
- Intervención en La Deficiencia MotoricaDocumento19 páginasIntervención en La Deficiencia MotoricaAna Maria GutierrezAún no hay calificaciones
- Inspección SUNAFILDocumento4 páginasInspección SUNAFILcarlos6navarro6madueAún no hay calificaciones
- Casos Clínicos Laso ResoltosDocumento61 páginasCasos Clínicos Laso ResoltosDima YingAún no hay calificaciones
- Manual Primeros AUX Mod 1Documento23 páginasManual Primeros AUX Mod 1Astrid Chichilota MuñozAún no hay calificaciones
- 1.2. Ej de Textos AlumnDocumento12 páginas1.2. Ej de Textos Alumnsalima benslaimanAún no hay calificaciones
- PSICOTERAPIA ManuelaDocumento11 páginasPSICOTERAPIA ManuelaKarla Sosa QuinteroAún no hay calificaciones
- Folleto SRDocumento1 páginaFolleto SRHector Mario Cruz Moncada100% (1)
- Capítulo II Del AseoDocumento3 páginasCapítulo II Del AseoREALGameAún no hay calificaciones
- HIPOPITUITARISMODocumento1 páginaHIPOPITUITARISMOAndryk ZazuetaAún no hay calificaciones
- Historia Clínica OrtodonciaDocumento12 páginasHistoria Clínica OrtodonciaMelody Fernandez GarciaAún no hay calificaciones
- Aceite de La Vida ListoDocumento2 páginasAceite de La Vida ListoDayana NicolleAún no hay calificaciones
- Renan Mapa ConceptualDocumento3 páginasRenan Mapa ConceptualRenán Hector Sánchez CernaAún no hay calificaciones
- Liquidos y Electrolitos en CirugiaDocumento46 páginasLiquidos y Electrolitos en CirugiaMilena LozanoAún no hay calificaciones
- 12 - Tomo XII-115-123Documento9 páginas12 - Tomo XII-115-123Zoe RaponiAún no hay calificaciones
- Análisis de La Situación Epidemiológica de La Nefropatía Diabética en El Perú y La Región Ica Al Año 2020Documento22 páginasAnálisis de La Situación Epidemiológica de La Nefropatía Diabética en El Perú y La Región Ica Al Año 2020Zulema Lengua ElíasAún no hay calificaciones
- 00 - Plan de Contingencia Limpieza y Desinfeccion (Covid 19) REV 0. 2020Documento13 páginas00 - Plan de Contingencia Limpieza y Desinfeccion (Covid 19) REV 0. 2020AntonioDeValenciaAún no hay calificaciones
- SEMANA 7 MICROBIOLOGIA - InformeDocumento25 páginasSEMANA 7 MICROBIOLOGIA - InformeGoose ZklAún no hay calificaciones
- De Mi Tío de AméricaDocumento4 páginasDe Mi Tío de AméricaAlejandro RozadoAún no hay calificaciones
- Higroma Quistico.Documento11 páginasHigroma Quistico.Lewis Oswaldo Yacila ZapataAún no hay calificaciones
- Plagas Industria AlimentariaDocumento8 páginasPlagas Industria AlimentariaSANDRA RAQUEL MORAAún no hay calificaciones
- Analisis Pelicula La Isla Siniestra 22Documento7 páginasAnalisis Pelicula La Isla Siniestra 22leonel galeano ibarra100% (1)
- Pgi 9 Programa de Salud OcupacionalDocumento27 páginasPgi 9 Programa de Salud OcupacionalYenny Carolina Hernandez GarnicaAún no hay calificaciones
- Inf Sobre La Iso 45000Documento11 páginasInf Sobre La Iso 45000leonelalbertodiaz78Aún no hay calificaciones
- 08 Estudio de ItuDocumento13 páginas08 Estudio de ItuErika Berenice Lopez RodriguezAún no hay calificaciones
- CodigosDocumento2 páginasCodigosVey HernandezAún no hay calificaciones
- Flashcards Vacunas Que Te Funcionaran Durante La Carrera - Flashcards-Vacunas-Que-Te-Funcionaran-Durante-La-CarreraDocumento8 páginasFlashcards Vacunas Que Te Funcionaran Durante La Carrera - Flashcards-Vacunas-Que-Te-Funcionaran-Durante-La-CarreraYairs JLAún no hay calificaciones
- Criaderos by J. FariñaDocumento7 páginasCriaderos by J. FariñaGrisetAún no hay calificaciones
- EPISTAXISDocumento45 páginasEPISTAXISJazmin Perez Lima QuirogaAún no hay calificaciones
- Tecnicas y Manejo de La Conducta Aplicada y Orientadas Al Higienista DentalDocumento20 páginasTecnicas y Manejo de La Conducta Aplicada y Orientadas Al Higienista DentalMaria RosseAún no hay calificaciones