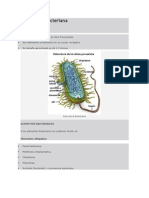
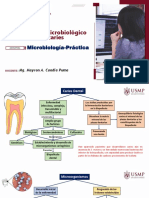
También podría gustarte
- Análisis funcional y microbiológico de derivados lácteos y cárnicosDe EverandAnálisis funcional y microbiológico de derivados lácteos y cárnicosAún no hay calificaciones
- Biología Molecular y Celular: Una guía introductoria para aprender Biología Celular y MolecularDe EverandBiología Molecular y Celular: Una guía introductoria para aprender Biología Celular y MolecularAún no hay calificaciones
- Clase 4 - Bacterias I ParteDocumento6 páginasClase 4 - Bacterias I ParteciinmimiAún no hay calificaciones
- Taller 3 FinalDocumento9 páginasTaller 3 FinalLuisAún no hay calificaciones
- Taller Morfología y Estructura BacterianaDocumento8 páginasTaller Morfología y Estructura BacterianaMelissa Tamayo GómezAún no hay calificaciones
- Estructuras esenciales y no esenciales de las bacteriasDocumento5 páginasEstructuras esenciales y no esenciales de las bacterias0511910049 EDINSON RAFAEL MEDINA MORON ESTUDIANTE ACTIVOAún no hay calificaciones
- 1203 BacteriasDocumento61 páginas1203 BacteriasCarlos ChávezAún no hay calificaciones
- UD2 - Aplicación de Técnicas de Tinción y Observación M.ODocumento15 páginasUD2 - Aplicación de Técnicas de Tinción y Observación M.ONoeliaAún no hay calificaciones
- Estructura bacteriana: características claveDocumento3 páginasEstructura bacteriana: características claveILSE YANET CRUZ DELGADOAún no hay calificaciones
- Caracteristicas Morfologicas de Las BacteriasDocumento9 páginasCaracteristicas Morfologicas de Las Bacteriaspollito138875% (4)
- Microbiologia y Parasitologia Guia 1Documento5 páginasMicrobiologia y Parasitologia Guia 1saraAún no hay calificaciones
- Informe Unidad I de Micro BiologiaDocumento21 páginasInforme Unidad I de Micro BiologiaelinaAún no hay calificaciones
- Anatomia Bacteriana.Documento6 páginasAnatomia Bacteriana.Rosmely Rojas50% (2)
- Bacterias - Método tinción GramDocumento18 páginasBacterias - Método tinción GramAntonio Bladimir Talledo TrellesAún no hay calificaciones
- Tincion GrammDocumento10 páginasTincion GrammdannyAún no hay calificaciones
- Estructura BacterianaDocumento11 páginasEstructura BacterianaCarlos Eduardo Celis Seclén0% (1)
- Taller MicrobiologiaDocumento5 páginasTaller MicrobiologiaKaren Ximena Cordoba OrdonezAún no hay calificaciones
- Bacteriología y MicrobiotaDocumento18 páginasBacteriología y MicrobiotaherlimarAún no hay calificaciones
- Estructuras celulares en Archaea y MoneraDocumento9 páginasEstructuras celulares en Archaea y MoneraAndry MartinezAún no hay calificaciones
- Estructura BacterianaDocumento3 páginasEstructura BacterianaDaniel CandAún no hay calificaciones
- Bacteria Gram PositivaDocumento35 páginasBacteria Gram PositivaMarcelino Duran Ruiz100% (1)
- Apuntes de MicrobiologíaDocumento23 páginasApuntes de MicrobiologíaHenz CarolAún no hay calificaciones
- Estructura de Las BacteriasDocumento5 páginasEstructura de Las BacteriasDulcesita Jackeline Gomez ZumaetaAún no hay calificaciones
- Estructura bacterianaDocumento8 páginasEstructura bacterianaJesus Rafael AstudilloAún no hay calificaciones
- Estructura de La Célula BacterianaDocumento3 páginasEstructura de La Célula BacterianawilmerAún no hay calificaciones
- Citología BacterianaDocumento12 páginasCitología BacterianaCarolina GalarzaAún no hay calificaciones
- Clase 3 Bacterias Morfologia EstructuraDocumento5 páginasClase 3 Bacterias Morfologia Estructuralesly quirozAún no hay calificaciones
- Estructura y morfología bacteriana enDocumento13 páginasEstructura y morfología bacteriana enTania Vazquez DornaAún no hay calificaciones
- Estructura BacterianaDocumento6 páginasEstructura BacterianaLuz Nayeli Caracara OrtizAún no hay calificaciones
- Reproducción BacterianaDocumento17 páginasReproducción BacterianaOriana Hnnd'z-mAún no hay calificaciones
- Unidad 3 - BacteriasDocumento19 páginasUnidad 3 - BacteriasDouglas R. MartínezAún no hay calificaciones
- Teoria MicrobiologiaDocumento54 páginasTeoria MicrobiologiaPedro Luis Gutierrez O.Aún no hay calificaciones
- Resumen Sordelli (Caps 1,2,3,5 y 6)Documento26 páginasResumen Sordelli (Caps 1,2,3,5 y 6)Ricardo AguileraAún no hay calificaciones
- Morfologia MicrobianaDocumento9 páginasMorfologia Microbiana.:("*"BLacK BuLLeT"*"):.33% (3)
- Estructura BacterianaDocumento7 páginasEstructura BacterianaMartha De La VegaAún no hay calificaciones
- Bacteriología: taxonomía, estructura y coloración de GramDocumento7 páginasBacteriología: taxonomía, estructura y coloración de GramanaAún no hay calificaciones
- Estructura Constante de La Célula BacterianaDocumento14 páginasEstructura Constante de La Célula BacterianaELDAAún no hay calificaciones
- Micro Guia EnteraDocumento30 páginasMicro Guia EnteraAlexia SosaAún no hay calificaciones
- Micro II Temas 1 y 2Documento18 páginasMicro II Temas 1 y 2herlimarAún no hay calificaciones
- Citologia Bacteriana 1 Clase 5sDocumento48 páginasCitologia Bacteriana 1 Clase 5sYurema CHAún no hay calificaciones
- Estructura bacteriana y morfologíaDocumento39 páginasEstructura bacteriana y morfologíaFernanda Vilches67% (3)
- Morfología y clasificación de bacteriasDocumento22 páginasMorfología y clasificación de bacteriasMaryam YousefAún no hay calificaciones
- Célula ProcariotaDocumento14 páginasCélula Procariota1111Aún no hay calificaciones
- Estructura y función de las células procariotasDocumento11 páginasEstructura y función de las células procariotasnicov04Aún no hay calificaciones
- TEMA 5 Bacterias MicrobiologiaDocumento12 páginasTEMA 5 Bacterias MicrobiologiaJulio LunaAún no hay calificaciones
- Generalidades BacteriologicasDocumento23 páginasGeneralidades BacteriologicasnicolasAún no hay calificaciones
- Bacteria ClasesDocumento23 páginasBacteria ClasesSalomonKingAún no hay calificaciones
- Microbiología e Inmunología FINAL. Gonzalez Luisiana Belén PDFDocumento86 páginasMicrobiología e Inmunología FINAL. Gonzalez Luisiana Belén PDFAgostina BlancoAún no hay calificaciones
- Bacteriología parcial 1: generalidades sobre morfología, estructura y clasificación de bacteriasDocumento11 páginasBacteriología parcial 1: generalidades sobre morfología, estructura y clasificación de bacteriasFelipe Garcia MartinezAún no hay calificaciones
- Estructura BacterianaDocumento6 páginasEstructura BacterianaDarlyn Valverde CernaAún no hay calificaciones
- Clase 4Documento22 páginasClase 4GenAún no hay calificaciones
- Genoma bacteriano: estructura y reproducciónDocumento6 páginasGenoma bacteriano: estructura y reproducciónEve Ivana Choque Gutierrez100% (1)
- Capitulo 26Documento9 páginasCapitulo 26Monse BeltranAún no hay calificaciones
- Bacterias FitopatologiaDocumento6 páginasBacterias FitopatologiaMelany BobadillaAún no hay calificaciones
- Taller de Repaso Morfologia MicrobianaDocumento20 páginasTaller de Repaso Morfologia MicrobianaJuliana RicoAún no hay calificaciones
- GlicocálixDocumento4 páginasGlicocálixSKEICY NICOLL NAVARRO OLIVERA100% (1)
- SMEAGOLDocumento24 páginasSMEAGOLAracely AjataAún no hay calificaciones
- Estruct Bacteria Apuntes PDFDocumento10 páginasEstruct Bacteria Apuntes PDFBioloscarAún no hay calificaciones
- ABO LibroDocumento19 páginasABO Librovalentina perezAún no hay calificaciones
- Bacteriología: Estructura y fisiología de las bacteriasDocumento39 páginasBacteriología: Estructura y fisiología de las bacteriasliz100% (1)
- Caracteristicas Generales de Los Microorganismos PROCARIOTAS SCBDocumento24 páginasCaracteristicas Generales de Los Microorganismos PROCARIOTAS SCBMarcela L. Garay BarbuyaniAún no hay calificaciones
- Si El Conocimiento Es Posible Como Se OriginaDocumento10 páginasSi El Conocimiento Es Posible Como Se OriginaMarcela L. Garay Barbuyani100% (1)
- Clase IV La Verdad Como Problema FilosóficoDocumento7 páginasClase IV La Verdad Como Problema FilosóficoMarcela L. Garay BarbuyaniAún no hay calificaciones
- Que Debo HacerDocumento7 páginasQue Debo HacerMarcela L. Garay Barbuyani100% (1)
- Clase III GnoseologíaDocumento7 páginasClase III GnoseologíaMarcela L. Garay BarbuyaniAún no hay calificaciones
- Caracteristicas Generales de Microorganismos Cont-Convertido para SCBDocumento5 páginasCaracteristicas Generales de Microorganismos Cont-Convertido para SCBMarcela L. Garay BarbuyaniAún no hay calificaciones
- Caracteristicas Generales de Microorganismos Cont-Convertido para SCBDocumento5 páginasCaracteristicas Generales de Microorganismos Cont-Convertido para SCBMarcela L. Garay BarbuyaniAún no hay calificaciones
- Garaymarcelaliliana Trabajopractico1 CapitalDocumento4 páginasGaraymarcelaliliana Trabajopractico1 CapitalMarcela L. Garay BarbuyaniAún no hay calificaciones
- Caracteristicas Generales de Microorganismos Cont-Convertido para SCBDocumento5 páginasCaracteristicas Generales de Microorganismos Cont-Convertido para SCBMarcela L. Garay BarbuyaniAún no hay calificaciones
- Para Que Ensen Amos BiologiaDocumento2 páginasPara Que Ensen Amos BiologiaMarcela L. Garay BarbuyaniAún no hay calificaciones
- Chaim Weizman0 ForoDocumento1 páginaChaim Weizman0 ForoMarcela L. Garay BarbuyaniAún no hay calificaciones
- Trabajo Laboratorio1Documento6 páginasTrabajo Laboratorio1Marcela L. Garay BarbuyaniAún no hay calificaciones
- La Llegada de Una Vacuna Frente Al SARSDocumento7 páginasLa Llegada de Una Vacuna Frente Al SARSMarcela L. Garay BarbuyaniAún no hay calificaciones
- Guia APADocumento11 páginasGuia APAUriel PerezAún no hay calificaciones
- Activ de Revision BiomoleculasDocumento1 páginaActiv de Revision BiomoleculasMarcela L. Garay BarbuyaniAún no hay calificaciones
- Alberdi y Sarmiento 2.2Documento3 páginasAlberdi y Sarmiento 2.2Marcela L. Garay BarbuyaniAún no hay calificaciones
- Para Que Ensen Amos BiologiaDocumento2 páginasPara Que Ensen Amos BiologiaMarcela L. Garay BarbuyaniAún no hay calificaciones
- Teoria Del AprendizajeDocumento32 páginasTeoria Del AprendizajeALEJANDROAún no hay calificaciones
- Evolucion03 100418065842 Phpapp02Documento29 páginasEvolucion03 100418065842 Phpapp02Maka Gomez Ibarra100% (2)
- Contreras Ensenanza Cap1Documento22 páginasContreras Ensenanza Cap1Maria Belen AcuñaAún no hay calificaciones
- Plantilla para Tesis Con Normas APA, Formato APADocumento18 páginasPlantilla para Tesis Con Normas APA, Formato APANazkterAún no hay calificaciones
- Tesis 1235 AnsiedadDocumento151 páginasTesis 1235 AnsiedadMarcela L. Garay BarbuyaniAún no hay calificaciones
- Contreras Ensenanza Cap1Documento22 páginasContreras Ensenanza Cap1Maria Belen AcuñaAún no hay calificaciones
- Zoología HickmanDocumento942 páginasZoología HickmanMarcela L. Garay Barbuyani92% (13)
- Calor y TemperaturaDocumento1 páginaCalor y TemperaturaMarcela L. Garay BarbuyaniAún no hay calificaciones
- 1 PteridofitasDocumento109 páginas1 PteridofitasHo SeokAún no hay calificaciones
- Organizacion de Las Celulas 1Documento15 páginasOrganizacion de Las Celulas 1Marcela L. Garay BarbuyaniAún no hay calificaciones
- CORTE GTRANSVERSAL DE UN TAlloDocumento3 páginasCORTE GTRANSVERSAL DE UN TAlloMarcela L. Garay BarbuyaniAún no hay calificaciones
- 10 Tallo Estruct PrimariaDocumento7 páginas10 Tallo Estruct Primariavodoochild_Ar80Aún no hay calificaciones
- GimnospermasDocumento20 páginasGimnospermasWilser DíazAún no hay calificaciones
- La Planta y Sus Partes para Segundo de PrimariaDocumento14 páginasLa Planta y Sus Partes para Segundo de PrimariaYurima PalenzuelaAún no hay calificaciones
- Trabajo Unidad 1Documento13 páginasTrabajo Unidad 1Sergio Luis Pacheco ColonAún no hay calificaciones
- Paper Proyecto HidropónicoDocumento3 páginasPaper Proyecto HidropónicoAlexanderTasinchanoAún no hay calificaciones
- VidDocumento9 páginasVidJuan Pablo Ricci RipamonttiAún no hay calificaciones
- Practica Botanica FrutoDocumento6 páginasPractica Botanica FrutoSilvia MartinezAún no hay calificaciones
- Index - 2022-11-10T084218.677Documento10 páginasIndex - 2022-11-10T084218.677KoiyzedAún no hay calificaciones
- Abono OrganicoDocumento8 páginasAbono OrganicoLizardo Garcia Alejandria100% (1)
- Mejoramiento Genético Del Ganado VacunoDocumento18 páginasMejoramiento Genético Del Ganado VacunoAriana DanielaAún no hay calificaciones
- Micología-Hongos y sus infeccionesDocumento4 páginasMicología-Hongos y sus infeccionesMILENE CHIRIBOGAAún no hay calificaciones
- Esperamatogenbesis en BovinosDocumento53 páginasEsperamatogenbesis en BovinosVALERY DANIELA VARELA MEJIAAún no hay calificaciones
- Programa de La Materia Biologia CBCDocumento2 páginasPrograma de La Materia Biologia CBCXimena RodriguezAún no hay calificaciones
- Ficha Seres Vivos CastellanoDocumento75 páginasFicha Seres Vivos Castellanolferrer1995Aún no hay calificaciones
- Identificación de Fauna NativaDocumento7 páginasIdentificación de Fauna NativaNath E VizueteAún no hay calificaciones
- Comprensión Lectora 3º EP-El OkapiDocumento6 páginasComprensión Lectora 3º EP-El Okapinmorenozudaire80Aún no hay calificaciones
- Célula Vegetal - Wikipedia, La Enciclopedia LibreDocumento37 páginasCélula Vegetal - Wikipedia, La Enciclopedia LibreKevin HanccoAún no hay calificaciones
- Biodiversidad EcológicaDocumento35 páginasBiodiversidad EcológicaJhoseline Jerly100% (1)
- Guia 4 NaturalesDocumento5 páginasGuia 4 NaturalesDiaz SharonAún no hay calificaciones
- Biologia de Los Organismos Celulares y Plantas CLASE 3Documento41 páginasBiologia de Los Organismos Celulares y Plantas CLASE 3Matias RolónAún no hay calificaciones
- Silabo: Universidad Nacional Del Callao Facultad Ciencias de La Salud Escuela Profesional de EnfermeríaDocumento12 páginasSilabo: Universidad Nacional Del Callao Facultad Ciencias de La Salud Escuela Profesional de Enfermeríaamerico Hancco salasAún no hay calificaciones
- La Secadera (En Pasifloras) Badea.Documento2 páginasLa Secadera (En Pasifloras) Badea.Reinaldo BautistaAún no hay calificaciones
- Práctica 5 ParasitologíaDocumento11 páginasPráctica 5 ParasitologíaAxelWarnerAún no hay calificaciones
- Aislamiento de ColoniasDocumento7 páginasAislamiento de Coloniaslissette1809Aún no hay calificaciones
- Alteraciones GeneticasDocumento15 páginasAlteraciones GeneticasJosue Boris Lipa SandovalAún no hay calificaciones
- Yemil - Evolución y AdaptaciónDocumento2 páginasYemil - Evolución y AdaptacióncristoferAún no hay calificaciones
- Diagnóstico microbiológico de la caries dentalDocumento15 páginasDiagnóstico microbiológico de la caries dentalaxel antonioAún no hay calificaciones
- Árboles de Gijón - FRESNODocumento2 páginasÁrboles de Gijón - FRESNOAntonio De Paz CabelloAún no hay calificaciones
- Programa Presupuestal de Enfermedades Metaxénicas y ZoonosisDocumento5 páginasPrograma Presupuestal de Enfermedades Metaxénicas y Zoonosisestudio lovon sanchezAún no hay calificaciones
- Reproducción Asexual (Tiptico)Documento2 páginasReproducción Asexual (Tiptico)Walter Dela CruzAún no hay calificaciones
- Taller No. 1. Biodiversidad Biotecnología Bioprospección y Ómicas VFDocumento8 páginasTaller No. 1. Biodiversidad Biotecnología Bioprospección y Ómicas VFJulian RengifoAún no hay calificaciones