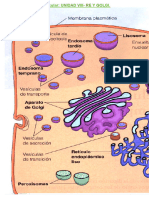
También podría gustarte
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012De EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012Aún no hay calificaciones
- Aparato de GolgiDocumento12 páginasAparato de GolgiAurelia Gonzales CalleAún no hay calificaciones
- Aparato de Golgi. LisosomasDocumento37 páginasAparato de Golgi. LisosomasAlexa RodríguezAún no hay calificaciones
- Aparato de GolgiDocumento34 páginasAparato de GolgiMoisés J. Duran CanevaroAún no hay calificaciones
- Aparato de GolgiDocumento4 páginasAparato de Golgipaz_saavedra_3100% (1)
- VII. Aparato de GolgiDocumento12 páginasVII. Aparato de GolgiKevin JoseAún no hay calificaciones
- VII. Aparato de GolgiDocumento12 páginasVII. Aparato de GolgiKevin JoseAún no hay calificaciones
- Aparato de GolgiDocumento12 páginasAparato de Golgiceciliaperez9751Aún no hay calificaciones
- Vacuolas JJJJJJJJJJJDocumento8 páginasVacuolas JJJJJJJJJJJmiguelon21Aún no hay calificaciones
- Aparato de GolgiDocumento4 páginasAparato de GolgiANDY MAURICIO YAGUANA SARITAMAAún no hay calificaciones
- El Aparato de Golgi Se Encarga de La Distribución y El Envío de Los Productos Químicos de La CélulaDocumento10 páginasEl Aparato de Golgi Se Encarga de La Distribución y El Envío de Los Productos Químicos de La CélulaN̛a͠g͞e̴r̕ M̕e͏d͠i͟na̛Aún no hay calificaciones
- Aparato de Golgi, Lisosomas y PeroxisomasDocumento46 páginasAparato de Golgi, Lisosomas y PeroxisomasYennifer Lizeth Ignación LeónAún no hay calificaciones
- Reti CuloDocumento4 páginasReti CuloJesusAún no hay calificaciones
- GolgiDocumento8 páginasGolgiAnnaAún no hay calificaciones
- GUIA DE RETÍCULO Y GOLGI TerminadaDocumento7 páginasGUIA DE RETÍCULO Y GOLGI TerminadaAndrea Ramás100% (5)
- Unidad 5 BioDocumento7 páginasUnidad 5 Biomolina.ailen99Aún no hay calificaciones
- Organelos PDFDocumento72 páginasOrganelos PDFVictor CarpioAún no hay calificaciones
- Sistema de EndomembranasDocumento35 páginasSistema de EndomembranasMaria Paula Quintero TamayoAún no hay calificaciones
- Sistema de EndomembranasDocumento4 páginasSistema de EndomembranasJosephGabrielAún no hay calificaciones
- Informe Re GolgiDocumento7 páginasInforme Re GolgienriqueAún no hay calificaciones
- García 5Documento14 páginasGarcía 5Guadalupe GarciaAún no hay calificaciones
- Re, Golgi, Lisosoma, Peroxisoma, MitocondriaDocumento118 páginasRe, Golgi, Lisosoma, Peroxisoma, MitocondriaJosselin Saavedra LópezAún no hay calificaciones
- Unidad 2Documento41 páginasUnidad 2Matias VillagranAún no hay calificaciones
- Taller Sistema de EndomenbranasDocumento8 páginasTaller Sistema de EndomenbranasSandra Milena Forigua CastiblancoAún no hay calificaciones
- Tema 5Documento9 páginasTema 5Irene Pulido SerranoAún no hay calificaciones
- Complejo de Golgi UsacDocumento21 páginasComplejo de Golgi UsacKatherin CanoAún no hay calificaciones
- Seminario Lisosomas CarpetaDocumento11 páginasSeminario Lisosomas CarpetaLourdesCorreaB100% (1)
- Guia Paiep GolgiDocumento11 páginasGuia Paiep GolgiSusy FeliAún no hay calificaciones
- LISOSOMASDocumento7 páginasLISOSOMASDIEGO FERNANDO SANCHEZ DAVANAún no hay calificaciones
- 20.retículo Endoplasmico RugosoDocumento22 páginas20.retículo Endoplasmico RugosoNadia SanchezAún no hay calificaciones
- Citología Parcial 2Documento58 páginasCitología Parcial 2Aroa SilvaAún no hay calificaciones
- Sistema de Endomembranas 363014 Downloable 1758018Documento9 páginasSistema de Endomembranas 363014 Downloable 1758018Dayana BarahonaAún no hay calificaciones
- Qué Es El Retículo Endoplasmático RugosoDocumento2 páginasQué Es El Retículo Endoplasmático RugosoLAURA MELISSA PEREZ CACERESAún no hay calificaciones
- Taller EndomembranasDocumento9 páginasTaller EndomembranasPatty Lopez0% (1)
- Lección Complejo de GolgiDocumento11 páginasLección Complejo de GolgiMPaz VelaAún no hay calificaciones
- Resumen Endomembranas 01Documento3 páginasResumen Endomembranas 01joaquinlugo47Aún no hay calificaciones
- Biología Re y GolgiDocumento6 páginasBiología Re y GolgiNabila InsfranAún no hay calificaciones
- POLISACARIDODocumento6 páginasPOLISACARIDOGino RodriguezAún no hay calificaciones
- SVC Biología 08Documento7 páginasSVC Biología 08Ptricia OliverosAún no hay calificaciones
- 8.sistemas Membranosos de La Célula EucariotaDocumento9 páginas8.sistemas Membranosos de La Célula EucariotaAntony VillamarAún no hay calificaciones
- Resolucion Guia 4 - Ayudantias DBIO1070Documento11 páginasResolucion Guia 4 - Ayudantias DBIO1070MarianaAún no hay calificaciones
- Aparato de Golgi Lisosomas y VacuolasDocumento43 páginasAparato de Golgi Lisosomas y VacuolasscribdenesimoAún no hay calificaciones
- Tema 6 AcabadoDocumento13 páginasTema 6 AcabadoMagdalena Oñate SerranoAún no hay calificaciones
- Taller de BiologiaDocumento10 páginasTaller de BiologiaYuveidys Chamorro VergaraAún no hay calificaciones
- Biologia Grupo 1Documento34 páginasBiologia Grupo 1ana maria galloza garciaAún no hay calificaciones
- PeroxisomaDocumento7 páginasPeroxisomaDelia Sirpa BautistaAún no hay calificaciones
- SEMDocumento11 páginasSEMjulissagonzalezzuritaAún no hay calificaciones
- Vesículas de Transporte o de TransiciónDocumento4 páginasVesículas de Transporte o de TransiciónÁlex Willan Acuña SilvaAún no hay calificaciones
- Taller 2. Biologia Celular y MolecularDocumento7 páginasTaller 2. Biologia Celular y MolecularMARIA LUCIA PIESCHACÓN CHUZCANOAún no hay calificaciones
- Biologia Celular Banco de Preguntas FinalDocumento14 páginasBiologia Celular Banco de Preguntas Finalcharit martinez ñomaAún no hay calificaciones
- Aparato de Golgi O9Documento7 páginasAparato de Golgi O9Rosenildo Dariz Dos Santos100% (1)
- Cont 3Documento3 páginasCont 3ABIGAIL ESPINOZAAún no hay calificaciones
- Resumen BiologiaDocumento4 páginasResumen BiologiaImanol VillenazAún no hay calificaciones
- MEMBRANA. ExposicionDocumento16 páginasMEMBRANA. ExposicionJuan PabloAún no hay calificaciones
- Sistema de EndomenbranasDocumento25 páginasSistema de Endomenbranaselmedico2013Aún no hay calificaciones
- Capítulo IV - Adhesión Celular, Uniones Celular y Matriz ExtracelularDocumento12 páginasCapítulo IV - Adhesión Celular, Uniones Celular y Matriz ExtracelularFederico Rivadeneira CornejoAún no hay calificaciones
- Aparato de GolgiDocumento30 páginasAparato de GolgiGeorge Robles Morales100% (1)
- Aparato de GolgiDocumento9 páginasAparato de GolgiSebastián TamayoAún no hay calificaciones
- 7-5 Heterosacaridoscomplejos GlicoproteinasDocumento26 páginas7-5 Heterosacaridoscomplejos GlicoproteinascristhianDRBAún no hay calificaciones
- Spotify Terminos y CondicionesDocumento32 páginasSpotify Terminos y CondicionesRenzoAún no hay calificaciones
- Simbologia Del Diagrama de FlujoDocumento7 páginasSimbologia Del Diagrama de FlujoCarlos SilupuAún no hay calificaciones
- GUIA No. 6 GRADOS 10 Y 11 ED. ARTISTICADocumento2 páginasGUIA No. 6 GRADOS 10 Y 11 ED. ARTISTICALimon AmargóAún no hay calificaciones
- Geometria de Subduccion de La Placa de Nazca y La Placa SudamericanaDocumento20 páginasGeometria de Subduccion de La Placa de Nazca y La Placa SudamericanaJuan Manuel Rojas Huayanay100% (1)
- Tema 3 Control de InventariosDocumento38 páginasTema 3 Control de InventariosMarco RamírezAún no hay calificaciones
- Cariotipo HumanoDocumento6 páginasCariotipo HumanoDaniel Marquiño Alonzo PazAún no hay calificaciones
- Contenidos Administracion de Personal - Final PDFDocumento62 páginasContenidos Administracion de Personal - Final PDFYuliana Alexandra Echeverri PinedaAún no hay calificaciones
- Pueblos Indígenas Edición San Luis PotosíDocumento14 páginasPueblos Indígenas Edición San Luis PotosíJaimicocamilAún no hay calificaciones
- Carta de EncargoDocumento6 páginasCarta de EncargoCamilo PerezAún no hay calificaciones
- T6 P1 Cab Fredy Ge2023Documento8 páginasT6 P1 Cab Fredy Ge2023F.Angel C. gomezAún no hay calificaciones
- Trastorno de La Conducta Alimentaria Reporte de LecturaDocumento4 páginasTrastorno de La Conducta Alimentaria Reporte de LecturaPaola GonzalezAún no hay calificaciones
- Carta A La Flip PDFDocumento2 páginasCarta A La Flip PDFZonaceroAún no hay calificaciones
- Xdoc - MX Informe NaturaDocumento10 páginasXdoc - MX Informe NaturaMiguel Angel Flores MoralesAún no hay calificaciones
- Trabajo Práctico N1 SeñasDocumento4 páginasTrabajo Práctico N1 SeñasAnabella SteimbrecherAún no hay calificaciones
- Administracion de Base de DatosDocumento10 páginasAdministracion de Base de DatosJuan José Ochoa CadilloAún no hay calificaciones
- Bibliografia PDFDocumento3 páginasBibliografia PDFKevin Aaron Rivero QuispeAún no hay calificaciones
- Carlos Aquí BBDocumento3 páginasCarlos Aquí BBCarlos AlexiAún no hay calificaciones
- GASTROENTEROLOGÍADocumento42 páginasGASTROENTEROLOGÍACindy CarranzaAún no hay calificaciones
- Alcohol IsopropilicoDocumento9 páginasAlcohol IsopropilicoJames CiezaAún no hay calificaciones
- Vallejos LH PDFDocumento136 páginasVallejos LH PDFJjamer Jjamer JjamerAún no hay calificaciones
- Plan de Estudios 2011 Educación BásicaDocumento13 páginasPlan de Estudios 2011 Educación BásicaEfrén GutierrezAún no hay calificaciones
- Clara Isabel Fernández Rodicio PDFDocumento9 páginasClara Isabel Fernández Rodicio PDFRoman IslasAún no hay calificaciones
- Articulo Macnamara CefalometriaDocumento34 páginasArticulo Macnamara CefalometriaOscar GarciaAún no hay calificaciones
- Proceso de Fabricacion de La Madera Plastica 1Documento2 páginasProceso de Fabricacion de La Madera Plastica 1Henry Camilo Díaz Layton76% (21)
- Libros SapiensalesDocumento5 páginasLibros SapiensalesSybranAún no hay calificaciones
- Tarea 6Documento7 páginasTarea 6Liz RiveraAún no hay calificaciones
- Directorio Gobernadores Regionales 22 de Enero 2020Documento7 páginasDirectorio Gobernadores Regionales 22 de Enero 2020LIZETAún no hay calificaciones
- Perro Salvaje de AfricaDocumento5 páginasPerro Salvaje de AfricalulamazzuferiAún no hay calificaciones
- Categorías y Formas VerbalesDocumento4 páginasCategorías y Formas VerbalesAngie CuellarAún no hay calificaciones
- Guias Claves de Cierre FiscalDocumento24 páginasGuias Claves de Cierre FiscalJose Alfredo BarriosAún no hay calificaciones