También podría gustarte
- Metabolismo GlugogenoDocumento23 páginasMetabolismo GlugogenomarialebatzAún no hay calificaciones
- Metabolismo de Hidratos de CarbonoDocumento9 páginasMetabolismo de Hidratos de CarbonoNinfa LópezAún no hay calificaciones
- GLUCONEOGENESIS y Metabolismo Del Glucogeno 1Documento16 páginasGLUCONEOGENESIS y Metabolismo Del Glucogeno 1Ximena CabreraAún no hay calificaciones
- Gluco GenoDocumento56 páginasGluco GenoEfrainEdmundoPagaloTayupandaAún no hay calificaciones
- Metabolismo Del GlucógenoDocumento10 páginasMetabolismo Del GlucógenoBerta González GómezAún no hay calificaciones
- Metabolismo Del GlucógenoDocumento24 páginasMetabolismo Del Glucógenotupapirico1645Aún no hay calificaciones
- Clase 11-FarmaciaDocumento21 páginasClase 11-FarmaciaJefry AguirreAún no hay calificaciones
- Exposición BioquímicaDocumento20 páginasExposición BioquímicaLizette Danae Marquez BelloAún no hay calificaciones
- Lectura 08 CarbohidratosDocumento13 páginasLectura 08 CarbohidratosVannya Cristobal LoayzaAún no hay calificaciones
- Metabolismo de Hidaratos de CarbonoDocumento7 páginasMetabolismo de Hidaratos de CarbonoAldi VillalbaAún no hay calificaciones
- Clase 11Documento23 páginasClase 11Dalila Torres IsuizaAún no hay calificaciones
- Metabolismo Del GlucógenoDocumento18 páginasMetabolismo Del GlucógenoBENJAMIN GABRIEL ANTONIO GUTIERREZ DUARTEAún no hay calificaciones
- Metabolismo Del Glucógeno en Los AnimalesDocumento11 páginasMetabolismo Del Glucógeno en Los AnimalesFátima Jerez Arroyo100% (1)
- 6 Glucogeno Lisis y GenesisDocumento6 páginas6 Glucogeno Lisis y GenesisPaola MichaelAún no hay calificaciones
- Metabolismo del glucógenoDocumento27 páginasMetabolismo del glucógenocarloshl73Aún no hay calificaciones
- Metabolismo Del GlucogenoDocumento23 páginasMetabolismo Del GlucogenoFeer RangelAún no hay calificaciones
- GLUCONEOGENESISDocumento82 páginasGLUCONEOGENESISMARIA CAMILA MARIN NIETOAún no hay calificaciones
- Taller 4. Metabolismo Del GlucógenoDocumento12 páginasTaller 4. Metabolismo Del GlucógenofranAún no hay calificaciones
- I-Metabolismo de Hidratos de CarbonoDocumento8 páginasI-Metabolismo de Hidratos de Carbonosara alvesAún no hay calificaciones
- Semana 11 - Teoria BioquimicaDocumento31 páginasSemana 11 - Teoria BioquimicaLuciana AronesAún no hay calificaciones
- Apuntes Del Metabolismo Del GlucógenoDocumento6 páginasApuntes Del Metabolismo Del GlucógenoMariana crystel Cabanillas SalcidoAún no hay calificaciones
- GlucogenesisDocumento35 páginasGlucogenesisAbel ChávezAún no hay calificaciones
- Tema18, Metabolismo Del GlucógenoDocumento7 páginasTema18, Metabolismo Del GlucógenoR Jireh CarryAún no hay calificaciones
- Glucogenolisis Glucogénesis Cetogénesis 5Documento45 páginasGlucogenolisis Glucogénesis Cetogénesis 5DianaOrdoñez100% (1)
- Informe de Glucogenolisis Uu12Documento24 páginasInforme de Glucogenolisis Uu12dieg0280% (1)
- Metabolismo Del GlucógenoDocumento10 páginasMetabolismo Del GlucógenobiologiaibericaAún no hay calificaciones
- Bioquímica de La Nutrición IIDocumento12 páginasBioquímica de La Nutrición IIMaria Candela DuarteAún no hay calificaciones
- Metabolismo glucógeno hígado músculoDocumento47 páginasMetabolismo glucógeno hígado músculounnik06Aún no hay calificaciones
- Química - Metabolismo de Hidratos de CarbonosDocumento9 páginasQuímica - Metabolismo de Hidratos de CarbonosMary AcostaAún no hay calificaciones
- Tema 2 de MFH III - 100758Documento4 páginasTema 2 de MFH III - 100758mercedes YanezAún no hay calificaciones
- Metabolismo de los hidratos de carbonoDocumento28 páginasMetabolismo de los hidratos de carbonoFranco FilippoliAún no hay calificaciones
- VivianaMartínez - Consulta3 - Glucogenog...Documento15 páginasVivianaMartínez - Consulta3 - Glucogenog...Viviana MartinezAún no hay calificaciones
- GlucogenesisDocumento34 páginasGlucogenesisDignoAún no hay calificaciones
- SX McardleDocumento12 páginasSX McardleYeison PelufoAún no hay calificaciones
- GLUCOGENOGENESISfDocumento38 páginasGLUCOGENOGENESISfjhoana ocandoAún no hay calificaciones
- Metabolismo de CarboidratosDocumento23 páginasMetabolismo de CarboidratosVerónica VelásquezAún no hay calificaciones
- Metabolismo Del Glucógeno y GlucogenolisisDocumento4 páginasMetabolismo Del Glucógeno y GlucogenolisisJoshua RichAún no hay calificaciones
- GLUCÓGENODocumento4 páginasGLUCÓGENOJazmin TurcuttoAún no hay calificaciones
- RepasoDocumento18 páginasRepasoLILIANA MONSERRAT GARCIA MALVAEZAún no hay calificaciones
- Metabolismo de GlucogenoDocumento31 páginasMetabolismo de GlucogenoFelipe LavínAún no hay calificaciones
- Resume NDocumento6 páginasResume NAngela Milagros Ramos CalderónAún no hay calificaciones
- Taller GlucolisisDocumento4 páginasTaller GlucolisisRoberto Jose Diaz CastroAún no hay calificaciones
- RESÚMENESDocumento5 páginasRESÚMENESMariel León BonifacioAún no hay calificaciones
- Gluco Neo GenesisDocumento27 páginasGluco Neo GenesisAndres Leonardo Leguizamon GarciaAún no hay calificaciones
- Presentación 10 GLUCONEOGENESISDocumento30 páginasPresentación 10 GLUCONEOGENESISE Clinton LMAún no hay calificaciones
- 5 Metabolismo Del GlucogenoDocumento6 páginas5 Metabolismo Del GlucogenoGladys Del Carmen Vásquez AranivaAún no hay calificaciones
- Resumen Fundamentos Prueba KathleenDocumento24 páginasResumen Fundamentos Prueba KathleenAngélicaAún no hay calificaciones
- GlucógenoDocumento3 páginasGlucógenojuliaAún no hay calificaciones
- SeminarioDocumento8 páginasSeminarioRama Castro MartinezAún no hay calificaciones
- Practica Sobre Aislamiento Del Glucogeno Hepatico y Obtencion de GlucosaDocumento14 páginasPractica Sobre Aislamiento Del Glucogeno Hepatico y Obtencion de GlucosaMargot Mejia Sanchez100% (1)
- Metabolismo de Los Hidratos de CarbonoDocumento4 páginasMetabolismo de Los Hidratos de CarbonoLara OviedoAún no hay calificaciones
- GlucogenesisDocumento7 páginasGlucogenesisJareth Caldera100% (3)
- Quimia BiologicaDocumento10 páginasQuimia BiologicaSharon AlvarezAún no hay calificaciones
- Tema 5 Metabolismo Del GlucogenoDocumento15 páginasTema 5 Metabolismo Del GlucogenoConeko :3Aún no hay calificaciones
- Gluconeogenesis 2Documento73 páginasGluconeogenesis 2eduardoAún no hay calificaciones
- Semana 05 - Metabolismo Del Glucogeno y Otros CarbohidratosDocumento29 páginasSemana 05 - Metabolismo Del Glucogeno y Otros CarbohidratosZuriel Umiña CelisAún no hay calificaciones
- Glucosa y glucógeno: metabolismo y funcionesDocumento79 páginasGlucosa y glucógeno: metabolismo y funcionesHillary Lens LensAún no hay calificaciones
- Glucogenòlisis ExposiciònDocumento6 páginasGlucogenòlisis ExposiciònPaola Andrea Diaz ContrerasAún no hay calificaciones
- Apuntes de Tercer Parcial de BioquímicaDocumento25 páginasApuntes de Tercer Parcial de BioquímicaFranklin Omar García RománAún no hay calificaciones
- La Fiesta de San Pedro y San PabloDocumento1 páginaLa Fiesta de San Pedro y San PabloJohn JuradoAún no hay calificaciones
- Ejercicios Combinadas BasiscosDocumento4 páginasEjercicios Combinadas BasiscosJohn JuradoAún no hay calificaciones
- Portafolio Virtual IiDocumento6 páginasPortafolio Virtual IiJohn JuradoAún no hay calificaciones
- Portafolio VirtualDocumento11 páginasPortafolio VirtualJohn JuradoAún no hay calificaciones
- CLASSROOMDocumento6 páginasCLASSROOMJohn JuradoAún no hay calificaciones
- Personal Social Ii 5°y6° Sem 29Documento9 páginasPersonal Social Ii 5°y6° Sem 29John JuradoAún no hay calificaciones
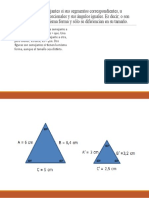
- Dos Figuras Son Semejantes Si Sus Segmentos CorrespondientesDocumento2 páginasDos Figuras Son Semejantes Si Sus Segmentos CorrespondientesJohn JuradoAún no hay calificaciones
- Buscar valor en rango Excel función BUSCARVDocumento9 páginasBuscar valor en rango Excel función BUSCARVJohn JuradoAún no hay calificaciones
- SMARTpro Convierte A Interfaz INteractivaDocumento22 páginasSMARTpro Convierte A Interfaz INteractivaJohn JuradoAún no hay calificaciones
- Cuando Llora Mi GuitarraDocumento1 páginaCuando Llora Mi GuitarraJohn Jurado100% (2)
- Funcion Buscarh en ExcelDocumento8 páginasFuncion Buscarh en ExcelJohn JuradoAún no hay calificaciones
- 2 Enseño Por Que Quiero A Ica 2022 FinalDocumento18 páginas2 Enseño Por Que Quiero A Ica 2022 FinalJohn JuradoAún no hay calificaciones
- Identifica Los LogosDocumento4 páginasIdentifica Los LogosJohn JuradoAún no hay calificaciones
- Portafolio VirtualDocumento11 páginasPortafolio VirtualJohn JuradoAún no hay calificaciones
- Google Classroom: Gestiona tu aula onlineDocumento6 páginasGoogle Classroom: Gestiona tu aula onlineJohn JuradoAún no hay calificaciones
- Autorizacion de Imagen y AudioDocumento1 páginaAutorizacion de Imagen y AudioJohn JuradoAún no hay calificaciones
- Bases Concurso de Canto 2021Documento5 páginasBases Concurso de Canto 2021John JuradoAún no hay calificaciones
- Trip TicoDocumento2 páginasTrip TicoJohn JuradoAún no hay calificaciones
- Ficha de InscripcionDocumento1 páginaFicha de InscripcionJohn Jurado100% (1)
- Crea mapas mentales gratis con MindomoDocumento18 páginasCrea mapas mentales gratis con MindomoLokito Maz NAAún no hay calificaciones
- FICHA DE MATEMATICA SEMANA 4 Dia 5Documento2 páginasFICHA DE MATEMATICA SEMANA 4 Dia 5John JuradoAún no hay calificaciones
- Fichas de Matematica Semana 5 AbrilDocumento5 páginasFichas de Matematica Semana 5 AbrilJohn JuradoAún no hay calificaciones
- Lab N 3Documento4 páginasLab N 3RAMIRO VILCA PEÑAAún no hay calificaciones
- Procedimiento para La Ondulacion 30-03-21Documento1 páginaProcedimiento para La Ondulacion 30-03-21RAUL LIGARDA MEDINAAún no hay calificaciones
- Problemas ReactoresDocumento11 páginasProblemas ReactoresJohnny GVAún no hay calificaciones
- Clase 7. Proceso de Disolución y Curvas de Solubilidad.Documento25 páginasClase 7. Proceso de Disolución y Curvas de Solubilidad.Ian RiscoAún no hay calificaciones
- Cap 1 Contminates AtmosfericosDocumento12 páginasCap 1 Contminates AtmosfericosSayonara TanquinaAún no hay calificaciones
- Sistema de Empaque y TrasabilidadDocumento43 páginasSistema de Empaque y TrasabilidadJannet Martínez purihuamanAún no hay calificaciones
- S05.s2 Material 2-Solucionario de Práctica Calificada 1 de Operaciones Unitarias-CGTDocumento12 páginasS05.s2 Material 2-Solucionario de Práctica Calificada 1 de Operaciones Unitarias-CGTmichelson buster manchego rodriguezAún no hay calificaciones
- PLIEGO DE ESPECIFICACIONES Ornamento Sidumss Norte Plan BDocumento11 páginasPLIEGO DE ESPECIFICACIONES Ornamento Sidumss Norte Plan BvladytechAún no hay calificaciones
- File 1459272403Documento10 páginasFile 1459272403DANIELA VANESSA HERMOZA SAAZAún no hay calificaciones
- 1.-Modulo 747 VictaulicDocumento2 páginas1.-Modulo 747 VictaulicOswaldo Saldivar LinaresAún no hay calificaciones
- PVA y PVOHDocumento9 páginasPVA y PVOHKenny Alvarez OcandoAún no hay calificaciones
- QuimicaDocumento17 páginasQuimicaDIEGO FERNANDO RAMÍREZ PLAZASAún no hay calificaciones
- 10 Ejemplos de Hidroxidos Con Nombre y Formula URGENTEDocumento6 páginas10 Ejemplos de Hidroxidos Con Nombre y Formula URGENTEAngel Gamboa Morales100% (2)
- MangoTirasAlmibarDocumento20 páginasMangoTirasAlmibarAlejandro Jesus Escobar JihuañaAún no hay calificaciones
- 20preguntas - 2P - Yomari LoorDocumento5 páginas20preguntas - 2P - Yomari LoorladyAún no hay calificaciones
- Historia de La TablaDocumento7 páginasHistoria de La TablaLeard Rafael Peralta MalaverAún no hay calificaciones
- Digrama de FasesDocumento4 páginasDigrama de FasesDAMARIS RUTH MAURICIO LASTRAAún no hay calificaciones
- Nte Inen 5Documento6 páginasNte Inen 5Pilozo SolisAún no hay calificaciones
- Recursos y reservas petrolerasDocumento24 páginasRecursos y reservas petrolerasSebastian Camilo MorenoAún no hay calificaciones
- Segundo Laboratorio (Motor Petter)Documento16 páginasSegundo Laboratorio (Motor Petter)Jorg Meza ParionaAún no hay calificaciones
- Ejercicios de OrtografíaDocumento3 páginasEjercicios de Ortografíaerika3017 Pachuca GarciaAún no hay calificaciones
- QIE-20Y-09 (TP - Enlace Lewis) AGDocumento4 páginasQIE-20Y-09 (TP - Enlace Lewis) AGArnold CruzAún no hay calificaciones
- Caracterizacion de Teca, Patula y AcaciaDocumento21 páginasCaracterizacion de Teca, Patula y AcaciaCristianAún no hay calificaciones
- Absorción de NutrientesDocumento6 páginasAbsorción de NutrientesKeren Gissell CruzAún no hay calificaciones
- Preguntas Tipo IcfesDocumento3 páginasPreguntas Tipo IcfesKarol HuerfanoAún no hay calificaciones
- Tabelas ÁguaDocumento50 páginasTabelas ÁguaBDCFAún no hay calificaciones
- Nte Inen 2034-1995 EsponjasDocumento8 páginasNte Inen 2034-1995 EsponjasPaola Andrea Contreras MartinezAún no hay calificaciones
- Guía para diluir ácido cítrico al 50% y 33Documento2 páginasGuía para diluir ácido cítrico al 50% y 33Omar AlconAún no hay calificaciones
- HS Li700Documento3 páginasHS Li700Alexis F.G.Aún no hay calificaciones
- Actividad de Aprendizaje 1 Evidencia Productos QuimicosDocumento31 páginasActividad de Aprendizaje 1 Evidencia Productos QuimicosMery MambuscayAún no hay calificaciones
- Principios básicos de bioquímica de los alimentosDe EverandPrincipios básicos de bioquímica de los alimentosCalificación: 4.5 de 5 estrellas4.5/5 (2)
- Bioquímica: metabolismo energético, conceptos y aplicaciónDe EverandBioquímica: metabolismo energético, conceptos y aplicaciónCalificación: 4 de 5 estrellas4/5 (7)
- Manual técnico de refrigerantesDe EverandManual técnico de refrigerantesCalificación: 4 de 5 estrellas4/5 (4)
- Fisiología renal y metabolismo hidrosalino: Segunda ediciónDe EverandFisiología renal y metabolismo hidrosalino: Segunda ediciónAún no hay calificaciones
- La tabla periódica de los elementos químicosDe EverandLa tabla periódica de los elementos químicosCalificación: 1 de 5 estrellas1/5 (1)
- Notas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesDe EverandNotas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesAún no hay calificaciones
- Mediciones y métodos de uso común en el laboratorio de QuímicaDe EverandMediciones y métodos de uso común en el laboratorio de QuímicaCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Sistema de mejora continua de la calidad en el laboratorio: Teoría y prácticaDe EverandSistema de mejora continua de la calidad en el laboratorio: Teoría y prácticaAún no hay calificaciones
- La química es la cuestión: El orden de la vida y el caos que llevamos encimaDe EverandLa química es la cuestión: El orden de la vida y el caos que llevamos encimaCalificación: 5 de 5 estrellas5/5 (5)
- Fundamentos de procesos químicosDe EverandFundamentos de procesos químicosCalificación: 5 de 5 estrellas5/5 (3)
- Fisiología comparada del medio internoDe EverandFisiología comparada del medio internoCalificación: 3 de 5 estrellas3/5 (1)
- Polímeros: Generalidades y tendencias de investigación en ColombiaDe EverandPolímeros: Generalidades y tendencias de investigación en ColombiaCalificación: 5 de 5 estrellas5/5 (14)
- Curso Práctico de Química GeneralDe EverandCurso Práctico de Química GeneralCalificación: 4 de 5 estrellas4/5 (4)
- Fisicoquímica II: Materia, electricidad y magnetismo. Fuerzas y camposDe EverandFisicoquímica II: Materia, electricidad y magnetismo. Fuerzas y camposAún no hay calificaciones
- El arte del descanso: Descubre el método para dormir bien y descansar mejorDe EverandEl arte del descanso: Descubre el método para dormir bien y descansar mejorCalificación: 4.5 de 5 estrellas4.5/5 (4)
- Inteligencia física: Aprende a ver el mundo con la mente de un físicoDe EverandInteligencia física: Aprende a ver el mundo con la mente de un físicoCalificación: 5 de 5 estrellas5/5 (41)
- Fundamentos de química aplicados a las ciencias de la saludDe EverandFundamentos de química aplicados a las ciencias de la saludAún no hay calificaciones
- Estructura atómica y enlace químicoDe EverandEstructura atómica y enlace químicoCalificación: 3 de 5 estrellas3/5 (2)
- La química al alcance de todosDe EverandLa química al alcance de todosCalificación: 1 de 5 estrellas1/5 (1)
- La dieta turbo de La Dama de los jugos: Pierda peso en poco tiempo y de forma saludableDe EverandLa dieta turbo de La Dama de los jugos: Pierda peso en poco tiempo y de forma saludableAún no hay calificaciones
- Problemas resueltos de Química Analítica CuantitativaDe EverandProblemas resueltos de Química Analítica CuantitativaCalificación: 5 de 5 estrellas5/5 (3)