También podría gustarte
- Diferencias Hombre Mujer: Descubre Los Últimos Hallazgos Científicos Sobre Las Diferencias Entre Mujeres Y HombresDe EverandDiferencias Hombre Mujer: Descubre Los Últimos Hallazgos Científicos Sobre Las Diferencias Entre Mujeres Y HombresAún no hay calificaciones
- DIFERENCIACION SEXUAL Clases.Documento54 páginasDIFERENCIACION SEXUAL Clases.Anonymous CSudLPP1Aún no hay calificaciones
- DIFERENCIACIóN SEXUALDocumento43 páginasDIFERENCIACIóN SEXUALRITA MARISOL GUANO TRUJILLOAún no hay calificaciones
- Estados Intersexuales IDocumento26 páginasEstados Intersexuales Ikatia ordoñez aldanaAún no hay calificaciones
- Sistema GenitalDocumento39 páginasSistema GenitalGabriela MoretaAún no hay calificaciones
- T10 Diferenciacion SexualDocumento45 páginasT10 Diferenciacion SexualElga TitoAún no hay calificaciones
- Embriología Del Aparato ReproductorDocumento83 páginasEmbriología Del Aparato ReproductorEdgardo Leal VillarroelAún no hay calificaciones
- Actividad Teorica Sistema Renal y GenitalDocumento59 páginasActividad Teorica Sistema Renal y GenitalFátima Patricia Hernández huertaAún no hay calificaciones
- Desarrollo Embrionario Y Diferenciacion Sexual: 1.1. SexoDocumento19 páginasDesarrollo Embrionario Y Diferenciacion Sexual: 1.1. SexojuanAún no hay calificaciones
- Res - Trastornos de Diferenciación SexualDocumento28 páginasRes - Trastornos de Diferenciación SexualYennyHerreraQuirozAún no hay calificaciones
- CAPÍTULO 9 - Gametogénesis y FecundaciónDocumento48 páginasCAPÍTULO 9 - Gametogénesis y FecundaciónSandra milena Diaz mercadoAún no hay calificaciones
- E17 - Desarrollo Del Aparato Genital 2017 123-136 WebDocumento14 páginasE17 - Desarrollo Del Aparato Genital 2017 123-136 WebMayjo CopiasAún no hay calificaciones
- GUIA - MANEJO - ADS - DSD - SEEP WalterDocumento77 páginasGUIA - MANEJO - ADS - DSD - SEEP WalterWalter RuizAún no hay calificaciones
- Histo Semana 2Documento20 páginasHisto Semana 2HeydiAún no hay calificaciones
- Trastornos de La Dif. SexualDocumento36 páginasTrastornos de La Dif. SexualLeticia Isabel Rojo GonzalezAún no hay calificaciones
- Hermafroditismo PDFDocumento9 páginasHermafroditismo PDFLuisa MatzkAún no hay calificaciones
- Determinación y Diferenciación Sexual 2023Documento35 páginasDeterminación y Diferenciación Sexual 2023Fabian RodasAún no hay calificaciones
- Pract Embrio #1Documento49 páginasPract Embrio #1Aka AkaAún no hay calificaciones
- Diferenciación SomatosexualDocumento6 páginasDiferenciación Somatosexualdaniela paredesAún no hay calificaciones
- Gametogenesis y Desarrollo de La Primera SemanaDocumento47 páginasGametogenesis y Desarrollo de La Primera SemanaGloria CristovaoAún no hay calificaciones
- Embriolog A Del Sistema Genital FemeninoDocumento14 páginasEmbriolog A Del Sistema Genital Femeninoalemartin000Aún no hay calificaciones
- Funciones de Los Sistemas Reproductores.Documento41 páginasFunciones de Los Sistemas Reproductores.LunaAún no hay calificaciones
- Cuadro Comparativo Del Desarrollo de La GónadaDocumento14 páginasCuadro Comparativo Del Desarrollo de La GónadaMiguel Angel Parra GarcíaAún no hay calificaciones
- ActPed2000 10Documento12 páginasActPed2000 10lauraAún no hay calificaciones
- Gametogenesis: EmbriologíaDocumento35 páginasGametogenesis: EmbriologíaBrayan CunoAún no hay calificaciones
- Trastornos de La Diferenciación Sexual y Amenorreas - José Augusto Fernández RodriguezDocumento24 páginasTrastornos de La Diferenciación Sexual y Amenorreas - José Augusto Fernández RodriguezBereNize SandovalAún no hay calificaciones
- Estrategias de Determinación Sexual - En.esDocumento13 páginasEstrategias de Determinación Sexual - En.esLina Marcela Reyes RivasAún no hay calificaciones
- Embriologia Genital FemeninaDocumento79 páginasEmbriologia Genital FemeninaGEDEON1Aún no hay calificaciones
- Embriologia General + GametogénesisDocumento47 páginasEmbriologia General + GametogénesisjfreitezAún no hay calificaciones
- Desarrollo CompletoDocumento12 páginasDesarrollo CompletoVeronica AlexandraAún no hay calificaciones
- SGyR CompletoDocumento231 páginasSGyR CompletoSofía Belén MartiranoAún no hay calificaciones
- Defectos Congénitos y MorfogénesisDocumento17 páginasDefectos Congénitos y Morfogénesisgiovanna ramosAún no hay calificaciones
- Determinación Del Sexo - EspañolDocumento13 páginasDeterminación Del Sexo - EspañolPamela TiqueAún no hay calificaciones
- Todo Sobre El SryDocumento16 páginasTodo Sobre El SryDuberGonzálezAún no hay calificaciones
- Trastornos Del Desarrollo SexualDocumento4 páginasTrastornos Del Desarrollo SexualGenesis SánchezAún no hay calificaciones
- 6 GametogénesisDocumento65 páginas6 GametogénesisMEDICINA VETERINARIA ZOOTECNISTAAún no hay calificaciones
- Trastornos SexualesFinaalDocumento30 páginasTrastornos SexualesFinaalÁngeles vidal rodriguezAún no hay calificaciones
- Resumen - Determinación y Diferenciación SexualDocumento6 páginasResumen - Determinación y Diferenciación SexualEnder ManAún no hay calificaciones
- Espermatogenesis 20230905 122609 0000Documento16 páginasEspermatogenesis 20230905 122609 0000Uriel HuertaAún no hay calificaciones
- Clase I. Sexualidad, Determinación y Diferenciacion SexualDocumento54 páginasClase I. Sexualidad, Determinación y Diferenciacion SexualDietry Lillo UribeAún no hay calificaciones
- 10-12. Reproducción y ToxicidadDocumento47 páginas10-12. Reproducción y Toxicidadjavier caicedoAún no hay calificaciones
- Trastornos Del Desarrollo SexualDocumento4 páginasTrastornos Del Desarrollo SexualCarmen BustillosAún no hay calificaciones
- 2..gametogenesis y Primera SemanaDocumento54 páginas2..gametogenesis y Primera SemanaIsamar CaicerAún no hay calificaciones
- Dimorfismo Sexual CerebralDocumento0 páginasDimorfismo Sexual CerebralCristian Gutierrez UlloaAún no hay calificaciones
- Informe de Estado IntersexualDocumento5 páginasInforme de Estado IntersexualRosali Sara Ccuno QuispeAún no hay calificaciones
- FECUNDACIONDocumento31 páginasFECUNDACIONCRISTINA RAZAAún no hay calificaciones
- Revista 1 2012 Pag 9 A 18Documento10 páginasRevista 1 2012 Pag 9 A 18Joel Mulato SanchezAún no hay calificaciones
- Determinación Médico Legal de Sexo - 2011Documento74 páginasDeterminación Médico Legal de Sexo - 2011Jorge SolarAún no hay calificaciones
- Defectos Congénitos y MorfogénesisDocumento17 páginasDefectos Congénitos y MorfogénesisElizabethAlegria0% (1)
- Genetica Del SexoDocumento7 páginasGenetica Del SexoSaul Adrian Mamani SanchezAún no hay calificaciones
- TP GametogenesisDocumento24 páginasTP GametogenesisDebo AldereteAún no hay calificaciones
- GametogenesisDocumento40 páginasGametogenesisRommel PorrasAún no hay calificaciones
- Resumen - Fisio Reproductiva (Berne y Levy)Documento20 páginasResumen - Fisio Reproductiva (Berne y Levy)Yara MartinezAún no hay calificaciones
- Clase 8Documento50 páginasClase 8Esteban BianchiAún no hay calificaciones
- Embriologia Del Aparato Genital FemeninoDocumento12 páginasEmbriologia Del Aparato Genital FemeninoJuli BetancourtAún no hay calificaciones
- Diferenciación GonadalDocumento8 páginasDiferenciación GonadalLily Losada EstradaAún no hay calificaciones
- Desarrollo Embrionario y Anomalías Congénitas Que Afectan A La Reproducción - Revista AsebirDocumento27 páginasDesarrollo Embrionario y Anomalías Congénitas Que Afectan A La Reproducción - Revista AsebirSunshine AmoreAún no hay calificaciones
- Eje HHG Desarrollo Puberal y EnvejecimientoDocumento50 páginasEje HHG Desarrollo Puberal y EnvejecimientoPablo Rios BricioAún no hay calificaciones
- Act - 3.3 - Kizimia - Fantini - Resumen. Sexo y CerebroDocumento6 páginasAct - 3.3 - Kizimia - Fantini - Resumen. Sexo y CerebroRafa FanAún no hay calificaciones
- Intersexo. 2022-2Documento72 páginasIntersexo. 2022-2Franchesca Reyes AbreuAún no hay calificaciones
- PROYECTO DE INVESTIGACIÒN TenevDocumento7 páginasPROYECTO DE INVESTIGACIÒN Tenevmaria del carmenAún no hay calificaciones
- 43 8 PBDocumento49 páginas43 8 PBmaria del carmenAún no hay calificaciones
- Las Áreas Verdes de La Costanera Contarán Con Sistemas de Riego Artificial - El LitoralDocumento2 páginasLas Áreas Verdes de La Costanera Contarán Con Sistemas de Riego Artificial - El Litoralmaria del carmenAún no hay calificaciones
- Tonalidad I Acústica y Música I Ruido Tonal I Svantek AcademyDocumento13 páginasTonalidad I Acústica y Música I Ruido Tonal I Svantek Academymaria del carmenAún no hay calificaciones
- 02 - BGR - MTJ ET Pliego de Bases y CondicionesDocumento28 páginas02 - BGR - MTJ ET Pliego de Bases y Condicionesmaria del carmenAún no hay calificaciones
- Las Enzimas HepaticasDocumento8 páginasLas Enzimas HepaticasFrank Félix Gómez De LeónAún no hay calificaciones
- 5 Electroforesis de Proteínas en Acetato de Celulosa Proteínas SéricasDocumento9 páginas5 Electroforesis de Proteínas en Acetato de Celulosa Proteínas SéricasKath RodríguezAún no hay calificaciones
- BiotecnologiaDocumento4 páginasBiotecnologiaLUIS ALEJANDRO PANTOJA VALLEJOAún no hay calificaciones
- Taller Sistema Endocrino 8°ab 29 de JulioDocumento8 páginasTaller Sistema Endocrino 8°ab 29 de JulioLuis SernaAún no hay calificaciones
- Sindrome de La Vaca GordaDocumento6 páginasSindrome de La Vaca GordaLea HimAún no hay calificaciones
- CX de OtorrinoDocumento40 páginasCX de OtorrinoBrayan HemirAún no hay calificaciones
- Trabajo de PsicobiologiaDocumento2 páginasTrabajo de Psicobiologiayuli andrea millan aguirreAún no hay calificaciones
- Cuidados de Enfermeria en PancreatitisDocumento9 páginasCuidados de Enfermeria en PancreatitisCarlosCeciAún no hay calificaciones
- Farmacologia de La InsulinaDocumento67 páginasFarmacologia de La InsulinaKaren Andrea Soto MillanAún no hay calificaciones
- Efectos Del Estado en Ayunas Frente Al Estado Alimentado Ejercicio Sobre El Rendimiento y Después Del Ejercicio MetabolismoDocumento7 páginasEfectos Del Estado en Ayunas Frente Al Estado Alimentado Ejercicio Sobre El Rendimiento y Después Del Ejercicio MetabolismoEsteban arceAún no hay calificaciones
- Práctica #2 PDFDocumento4 páginasPráctica #2 PDFSantiago Perez OlveraAún no hay calificaciones
- Hepatopatia CronicaDocumento28 páginasHepatopatia Cronicamauricio eduardo rojasAún no hay calificaciones
- 1794853-AF8.-Sistema EndócrinoDocumento4 páginas1794853-AF8.-Sistema EndócrinoCrisAún no hay calificaciones
- Anestesicos LocalesDocumento13 páginasAnestesicos LocalesAndres Perez HettingaAún no hay calificaciones
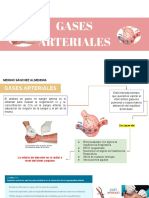
- Gases ArterialesDocumento23 páginasGases ArterialesLuz Ledesma OropezaAún no hay calificaciones
- Catabolismo de ProteínasDocumento2 páginasCatabolismo de ProteínasJose lorduyAún no hay calificaciones
- CLASE 3 Metabolismo Del SodioDocumento38 páginasCLASE 3 Metabolismo Del SodioValeria Tovar MartinezAún no hay calificaciones
- Tema 4.1 Nutrientes y AlimentaciónDocumento6 páginasTema 4.1 Nutrientes y AlimentaciónNatalia Grimaldi SánchezAún no hay calificaciones
- El Sistema EndocrinoDocumento10 páginasEl Sistema Endocrinomelany pilozoAún no hay calificaciones
- Cómo Se Relacionan Los Chakras Con Tus Estados EmocionalesDocumento38 páginasCómo Se Relacionan Los Chakras Con Tus Estados EmocionalesLaima BruxAún no hay calificaciones
- Catalogo InBody 2017Documento16 páginasCatalogo InBody 2017JOCELYN ACEVEDOAún no hay calificaciones
- Reproductor FemeninoDocumento55 páginasReproductor FemeninoJulissa VizaAún no hay calificaciones
- 200 Preguntas Sobre Cámara HiperbáricaDocumento95 páginas200 Preguntas Sobre Cámara HiperbáricaEdwin Mondragón100% (3)
- Listado de Categorias de EnfermedadesDocumento8 páginasListado de Categorias de EnfermedadesSifu Hung ChileAún no hay calificaciones
- Formato ClinicoDocumento8 páginasFormato ClinicoLionel HillAún no hay calificaciones
- Tipos de Test Caseros de Embarazo Muy Fáciles de RealizarDocumento2 páginasTipos de Test Caseros de Embarazo Muy Fáciles de RealizarAlvarado YeniferAún no hay calificaciones
- PUEMR3Documento6 páginasPUEMR3Javier Burgos Cárdenas100% (1)
- Lo Invitamos A Conocer Master KeyDocumento1 páginaLo Invitamos A Conocer Master KeyZuleika VSAún no hay calificaciones
- OPINIÓN DOCUMENTADA DE LA ACTIVIDAD REALIZADA Endocrinología 2.8Documento2 páginasOPINIÓN DOCUMENTADA DE LA ACTIVIDAD REALIZADA Endocrinología 2.8Jorge Andre Clavijo ColonaAún no hay calificaciones
- Documento (3) - 1Documento8 páginasDocumento (3) - 1Efrain EncarnacionAún no hay calificaciones
- Sesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesDe EverandSesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesCalificación: 4.5 de 5 estrellas4.5/5 (13)
- Anatomía del caballo: Guía práctica ilustradaDe EverandAnatomía del caballo: Guía práctica ilustradaCalificación: 4 de 5 estrellas4/5 (9)
- Liberación del trauma: Perdón y temblor es el caminoDe EverandLiberación del trauma: Perdón y temblor es el caminoCalificación: 4 de 5 estrellas4/5 (4)
- Homo antecessor: El nacimiento de una especieDe EverandHomo antecessor: El nacimiento de una especieCalificación: 5 de 5 estrellas5/5 (1)
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- Historia del cuerpo humano: Evolución, salud y enfermedadDe EverandHistoria del cuerpo humano: Evolución, salud y enfermedadAún no hay calificaciones
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- Cuerpo Tóxico: Como Liberar Tu Cuerpo De Las Toxinas Externas E Internas, Y Evitar Asi Los Efectos De Los Radicales LibresDe EverandCuerpo Tóxico: Como Liberar Tu Cuerpo De Las Toxinas Externas E Internas, Y Evitar Asi Los Efectos De Los Radicales LibresCalificación: 5 de 5 estrellas5/5 (2)
- Cerebro y silencio: Las claves de la creatividad y la serenidadDe EverandCerebro y silencio: Las claves de la creatividad y la serenidadCalificación: 5 de 5 estrellas5/5 (2)
- GuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisDe EverandGuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisCalificación: 4 de 5 estrellas4/5 (9)
- Cerebros rotos: Pacientes asombrosos que me enseñaron a vivirDe EverandCerebros rotos: Pacientes asombrosos que me enseñaron a vivirCalificación: 5 de 5 estrellas5/5 (3)
- Inteligencia artificial: Una exploración filosófica sobre el futuro de la mente y la concienciaDe EverandInteligencia artificial: Una exploración filosófica sobre el futuro de la mente y la concienciaAna Isabel Sánchez DíezCalificación: 4 de 5 estrellas4/5 (3)
- La increíble conexión intestino cerebro: Descubre la relación entre las emociones y el equilibrio intestinalDe EverandLa increíble conexión intestino cerebro: Descubre la relación entre las emociones y el equilibrio intestinalCalificación: 4.5 de 5 estrellas4.5/5 (103)
- 200 tareas en terapia breve: 2ª ediciónDe Everand200 tareas en terapia breve: 2ª ediciónCalificación: 4.5 de 5 estrellas4.5/5 (33)
- El cerebro del niño explicado a los padresDe EverandEl cerebro del niño explicado a los padresCalificación: 4.5 de 5 estrellas4.5/5 (147)
- Psicología forense: Estudio de la mente criminalDe EverandPsicología forense: Estudio de la mente criminalCalificación: 4.5 de 5 estrellas4.5/5 (18)
- Manual de osteología de cráneo bovino y equinoDe EverandManual de osteología de cráneo bovino y equinoCalificación: 4.5 de 5 estrellas4.5/5 (7)
- PNL: Domina tu mente y aprende como atraer el dinero con técnicas de Programación NeurolingüísticaDe EverandPNL: Domina tu mente y aprende como atraer el dinero con técnicas de Programación NeurolingüísticaCalificación: 4.5 de 5 estrellas4.5/5 (34)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Diseño, construcción y pruebas de horno prototipo semicontinuo para producir cerámicaDe EverandDiseño, construcción y pruebas de horno prototipo semicontinuo para producir cerámicaAún no hay calificaciones
- Las Enfermedades comienzan y terminan en tu mente: Una guía para la autosanaciónDe EverandLas Enfermedades comienzan y terminan en tu mente: Una guía para la autosanaciónCalificación: 4 de 5 estrellas4/5 (4)
- UF0085 - Diagnóstico estético facial y corporalDe EverandUF0085 - Diagnóstico estético facial y corporalCalificación: 4 de 5 estrellas4/5 (5)
- Genética general: Libro de textoDe EverandGenética general: Libro de textoCalificación: 4.5 de 5 estrellas4.5/5 (11)