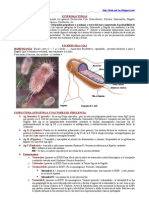
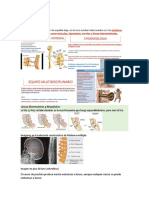
También podría gustarte
- Vibrio Cap 26Documento4 páginasVibrio Cap 26Maria GalgueraAún no hay calificaciones
- Unidad #5 MicrobiologiaDocumento11 páginasUnidad #5 MicrobiologiaErika GrabovieskiAún no hay calificaciones
- Resumen de Vibrio1Documento3 páginasResumen de Vibrio1Yulissa PittyAún no hay calificaciones
- Vibrio, Aeromonas y PlesiomonasDocumento8 páginasVibrio, Aeromonas y PlesiomonasFARID URIBE VALENCIAAún no hay calificaciones
- Vibrio CholeraeDocumento7 páginasVibrio CholeraeJan Negrete100% (1)
- Vibrio Campylobacter y HelicobacterDocumento11 páginasVibrio Campylobacter y HelicobacterEliza ChanAún no hay calificaciones
- 15 VibrioDocumento68 páginas15 VibriojuanAún no hay calificaciones
- Vibrio Parahaemolyticus-CholeraeDocumento8 páginasVibrio Parahaemolyticus-CholeraeNatalia Ramos MezaAún no hay calificaciones
- Bacilos Gram NegativosDocumento63 páginasBacilos Gram NegativosPaola Yanara VargasAún no hay calificaciones
- 6-2 Fam VibrionaceaeDocumento78 páginas6-2 Fam VibrionaceaelinurxerAún no hay calificaciones
- U2T7 - Género Vibrio, Género Campylobacter y Género HelicobacterDocumento26 páginasU2T7 - Género Vibrio, Género Campylobacter y Género HelicobacterLeonella UtreraAún no hay calificaciones
- Campylobacter, Helicobacter y VibrioDocumento6 páginasCampylobacter, Helicobacter y VibrioJeffrey LoayzaAún no hay calificaciones
- Vibrios ColeraDocumento72 páginasVibrios ColeraVictor Gavidia CaceresAún no hay calificaciones
- Género VibrioDocumento47 páginasGénero VibrioCristopherJairEstupiñanVirú100% (2)
- 13 - Vibrio CholeraeDocumento2 páginas13 - Vibrio CholeraealexAún no hay calificaciones
- Tema 37 VibrioDocumento7 páginasTema 37 VibrioStefany Graciela Hernandez RodriguezAún no hay calificaciones
- Vibrio AeromonasDocumento21 páginasVibrio AeromonasElizabeth SantillanAún no hay calificaciones
- CAMPYLOBACTERDocumento6 páginasCAMPYLOBACTERDahiana ViverosAún no hay calificaciones
- Exposición de VibrioDocumento20 páginasExposición de VibrioMateo GuerraAún no hay calificaciones
- VibriosDocumento2 páginasVibriosAmy Suazo SánchezAún no hay calificaciones
- Micro Sanitaria Tema 3.1Documento14 páginasMicro Sanitaria Tema 3.1AliciaAún no hay calificaciones
- Familia VibrionaceaeDocumento3 páginasFamilia VibrionaceaeJordi Canales100% (1)
- ColeraaaaDocumento20 páginasColeraaaaAle ChiapperoAún no hay calificaciones
- APUNTE Vibrio CholeraeDocumento2 páginasAPUNTE Vibrio CholeraeDiana BurbanoAún no hay calificaciones
- BIO150 Clase 13-Gram Negativo 2012 VIPDocumento77 páginasBIO150 Clase 13-Gram Negativo 2012 VIPDaniela Navarro SepúlvedaAún no hay calificaciones
- Diapositivas Vibrio Cholerae Sep Tiembre 2022Documento57 páginasDiapositivas Vibrio Cholerae Sep Tiembre 2022Luz AngelaAún no hay calificaciones
- Vibrio CholeraeDocumento2 páginasVibrio CholeraeXimena GarciaAún no hay calificaciones
- AeromonasDocumento4 páginasAeromonasPaola Daniela Quisbert SacariasAún no hay calificaciones
- 11 Sindrome Diarreico NeonatalDocumento15 páginas11 Sindrome Diarreico NeonatalJesus ChirinosAún no hay calificaciones
- Aeromonas y Plesiomonas WoedDocumento20 páginasAeromonas y Plesiomonas WoedAndrésAún no hay calificaciones
- E. Coli y Otras EnterobacteriasDocumento8 páginasE. Coli y Otras EnterobacteriasJosé Gabriel Quiroga VillagraAún no hay calificaciones
- Cap 28 Camplylobacter y HelycobacterDocumento4 páginasCap 28 Camplylobacter y HelycobacterMaria GalgueraAún no hay calificaciones
- Tema 8. Diarre Aguda. Infecciones de Transmisión SexualDocumento12 páginasTema 8. Diarre Aguda. Infecciones de Transmisión SexualGabriel PLaoAún no hay calificaciones
- Vibrio CholeraeDocumento4 páginasVibrio CholeraeDanya Astudillo VillalobosAún no hay calificaciones
- Informe Teoria 8Documento5 páginasInforme Teoria 8NICOLE ALEXANDRA ANICETO LLAURYAún no hay calificaciones
- Enfermedades Infecciosas IntestinalesDocumento14 páginasEnfermedades Infecciosas IntestinalesRolando RiosAún no hay calificaciones
- Infecciones Del Tracto Digestivo Estudiantes PDFDocumento28 páginasInfecciones Del Tracto Digestivo Estudiantes PDFlordgimli77Aún no hay calificaciones
- Ficha Vibrio CholoreaeDocumento3 páginasFicha Vibrio CholoreaeMarjorie Iveth Leyton OyarzoAún no hay calificaciones
- Trabajo Final Laboratorio EpidemiologiaDocumento9 páginasTrabajo Final Laboratorio EpidemiologiaIdalfi Mármol Pérez100% (1)
- 9 Teoria MicrobiologiaDocumento27 páginas9 Teoria Microbiologiawillian idrogo megoAún no hay calificaciones
- Vibrio CholeraeDocumento4 páginasVibrio CholeraeDamaris LancachoAún no hay calificaciones
- Ensayo Microobiologia y ParasitologiaDocumento14 páginasEnsayo Microobiologia y Parasitologia20440337Aún no hay calificaciones
- UntitledDocumento55 páginasUntitledEmilia Davila LazoAún no hay calificaciones
- 15 VibrioDocumento56 páginas15 Vibrioines pachecoAún no hay calificaciones
- VibRIO y Aeromonas (Cap 28)Documento8 páginasVibRIO y Aeromonas (Cap 28)David JuarezAún no hay calificaciones
- Microorganismos Utilizados Como Indicadores de Contaminación FecalDocumento20 páginasMicroorganismos Utilizados Como Indicadores de Contaminación FecalMargarita Agudelo RicoAún no hay calificaciones
- Familia VibrionaceaeDocumento25 páginasFamilia VibrionaceaeTatiana100% (1)
- Triptico VibrioDocumento2 páginasTriptico VibrioCristopherJairEstupiñanVirúAún no hay calificaciones
- Enfermedades Bacterianas en PecesDocumento39 páginasEnfermedades Bacterianas en PecesBrenda Fabiola ChavestaAún no hay calificaciones
- Bacilos Gramnegativos EntéricosDocumento7 páginasBacilos Gramnegativos EntéricosDIANA CAROLINA HERNANDEZ RIOSAún no hay calificaciones
- Fiebre Tifoidea, Shigelosis, ColeraDocumento9 páginasFiebre Tifoidea, Shigelosis, ColeraJesús ChacónAún no hay calificaciones
- E ColiDocumento5 páginasE ColiAngel SaldañaAún no hay calificaciones
- InfeccionesDocumento6 páginasInfeccionesLuis FCAún no hay calificaciones
- E Coli 2Documento30 páginasE Coli 2UnaTalLeyakziAún no hay calificaciones
- Vibrio Cholerae 2Documento5 páginasVibrio Cholerae 2vanessaAún no hay calificaciones
- H. PyloriDocumento9 páginasH. Pyloriclalisjuli956095Aún no hay calificaciones
- BacteriasDocumento3 páginasBacteriaserika velozAún no hay calificaciones
- EnterobacteriasDocumento52 páginasEnterobacteriasPeter RGAún no hay calificaciones
- Practica 2: Analisis Microbiologico de BebidasDocumento25 páginasPractica 2: Analisis Microbiologico de BebidasL C88% (16)
- Los bacteriófagos: Los virus que combaten infeccionesDe EverandLos bacteriófagos: Los virus que combaten infeccionesAún no hay calificaciones
- Ejercicio 14 Ciencias de La VidaDocumento4 páginasEjercicio 14 Ciencias de La VidaKarla Chucuan LeyvaAún no hay calificaciones
- Temas de Exposición Adulto Mayor 2022-2Documento3 páginasTemas de Exposición Adulto Mayor 2022-2Aracelly Rocio Bringas OcampoAún no hay calificaciones
- Diapositivas Enfermedades Prevalentes de La InfanciaDocumento29 páginasDiapositivas Enfermedades Prevalentes de La Infanciaonluheda85Aún no hay calificaciones
- Cadenas EpidemiologicasDocumento11 páginasCadenas EpidemiologicasJose OvandoAún no hay calificaciones
- Insectos VenenososDocumento4 páginasInsectos VenenososMoreli Emilia Ramírez SorianoAún no hay calificaciones
- Intoxicación Por Metales PesadosDocumento203 páginasIntoxicación Por Metales PesadosClauMcClauAún no hay calificaciones
- LumbalgiaDocumento18 páginasLumbalgiaMagui urrutiaAún no hay calificaciones
- Control de Artrópodos y Roedores de Importancia Sanitaria MS TareaDocumento3 páginasControl de Artrópodos y Roedores de Importancia Sanitaria MS TareaRosbelin Pichardo100% (2)
- Infecciones de Vías UrinariasDocumento22 páginasInfecciones de Vías Urinariasfatima alvaradoAún no hay calificaciones
- Certificado Medico 6256442Documento1 páginaCertificado Medico 6256442Emily TrávezAún no hay calificaciones
- 11 NeurologiaDocumento7 páginas11 Neurologiamir-medicinaAún no hay calificaciones
- Neutropenia y SX Hipereosinofilico - HEMATO (Autoguardado)Documento40 páginasNeutropenia y SX Hipereosinofilico - HEMATO (Autoguardado)Angelica GamezAún no hay calificaciones
- TetraciclinasDocumento37 páginasTetraciclinasMaria Pia RuizAún no hay calificaciones
- 03 Enfermedad Pelvica InflamatoriaDocumento25 páginas03 Enfermedad Pelvica InflamatoriaMalcon Enrique Alvarado CalderwoodAún no hay calificaciones
- Cuadro de GastroenterologíaDocumento31 páginasCuadro de GastroenterologíaMUNDO DISEÑOAún no hay calificaciones
- 252-Texto Del Artículo-267-1-10-20190108Documento6 páginas252-Texto Del Artículo-267-1-10-20190108gleinysAún no hay calificaciones
- Microbiologia y Parasitologia AlbumDocumento98 páginasMicrobiologia y Parasitologia AlbumMaricela RVAún no hay calificaciones
- Cuadro ComparativoDocumento5 páginasCuadro ComparativoJHOEL ALEX CLAROS GUZMANAún no hay calificaciones
- Estudio de Caso Charagua Sesiones 1-4Documento11 páginasEstudio de Caso Charagua Sesiones 1-4Celio CasanovaAún no hay calificaciones
- Gustavo JaraDocumento2 páginasGustavo JaraDarliana Ospina DuarteAún no hay calificaciones
- Fisterra Examenes de LaboratorioDocumento19 páginasFisterra Examenes de LaboratorioGaby GarridoAún no hay calificaciones
- 6.2.3 Laboratorios de Nivel IntermedioDocumento14 páginas6.2.3 Laboratorios de Nivel IntermedioEdithAún no hay calificaciones
- Patología Glandula TiroidesDocumento27 páginasPatología Glandula TiroidesEmmanuel VelazquezAún no hay calificaciones
- Tema 2 Etiología de Los Trastornos NeuropsicológicosDocumento21 páginasTema 2 Etiología de Los Trastornos NeuropsicológicosMarta Romero LópezAún no hay calificaciones
- TendiolDocumento1 páginaTendiolEdwin QuizhpiAún no hay calificaciones
- 2ML-FR-0002 Pliego de AntecedentesDocumento3 páginas2ML-FR-0002 Pliego de AntecedentesWilson PovedaAún no hay calificaciones
- Ficha Educacion Fisica-2024Documento2 páginasFicha Educacion Fisica-2024El Negrito LibreriaAún no hay calificaciones
- Pseudomonas, Legionella, Haemophilus, Brucella, Francisella y PasteurellaDocumento46 páginasPseudomonas, Legionella, Haemophilus, Brucella, Francisella y Pasteurellakarla saucedoAún no hay calificaciones
- Esquema de VacunasDocumento19 páginasEsquema de Vacunasricardo aguilarAún no hay calificaciones
- Mapa Mental DiarreaDocumento4 páginasMapa Mental DiarreaJoel GuishaAún no hay calificaciones