También podría gustarte
- Cátedra: Greco: Introducción Ala EconomíaDocumento110 páginasCátedra: Greco: Introducción Ala EconomíaBrisa SaucedoAún no hay calificaciones
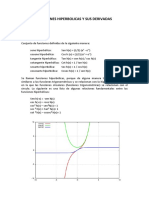
- Funciones Hiperbolicas y Sus DerivadasDocumento10 páginasFunciones Hiperbolicas y Sus DerivadasannaAún no hay calificaciones
- Apuntes GenomasurDocumento67 páginasApuntes GenomasurJuan José VasquezAún no hay calificaciones
- La sociología como ciencia: sus orígenes y característicasDocumento13 páginasLa sociología como ciencia: sus orígenes y característicasjuansartiAún no hay calificaciones
- Programa EX ZERBA 2023Documento11 páginasPrograma EX ZERBA 2023Ann WasingerAún no hay calificaciones
- Resumen Primer Parcial Historia de La PsicologiaDocumento21 páginasResumen Primer Parcial Historia de La PsicologiaCamila AttentoAún no hay calificaciones
- NM - Geografia2 - America Latina PDFDocumento225 páginasNM - Geografia2 - America Latina PDFEmilia PolettiAún no hay calificaciones
- Guía Semiología SaabDocumento4 páginasGuía Semiología SaabLyane MatosAún no hay calificaciones
- Resumen para El Primer Parcial de ICSEDocumento12 páginasResumen para El Primer Parcial de ICSEmatias mariscalAún no hay calificaciones
- CLaudio Lopez Tarea 2Documento4 páginasCLaudio Lopez Tarea 2ClaudioAún no hay calificaciones
- Cuadernillo 1er Cuat 2024Documento79 páginasCuadernillo 1er Cuat 2024Loreley MartinezAún no hay calificaciones
- ICSE B Tema 1 - Valdez - Intensivo Diciembre 22Documento4 páginasICSE B Tema 1 - Valdez - Intensivo Diciembre 22Davi OliveiraAún no hay calificaciones
- Libro Unlz MetodologiaDocumento233 páginasLibro Unlz MetodologiaDailan Kifki100% (1)
- Resúmen Completo BIOLOGÍA CELULAR 54Documento115 páginasResúmen Completo BIOLOGÍA CELULAR 54diazrojaskiaraAún no hay calificaciones
- Hidrodinamica 2Documento13 páginasHidrodinamica 2Erika Huanqui CoilaAún no hay calificaciones
- Ipc Catedra Arroche KetzelmanDocumento12 páginasIpc Catedra Arroche Ketzelmanlolo roloAún no hay calificaciones
- El Valle de La Luna PDFDocumento386 páginasEl Valle de La Luna PDFPatricia OcampoAún no hay calificaciones
- Ejemplos de Los Casos de Integrales TrigonometricasDocumento5 páginasEjemplos de Los Casos de Integrales TrigonometricasmarjhoanAún no hay calificaciones
- Guia Del Capitulo 1 Resuelto Ipc FlaxDocumento6 páginasGuia Del Capitulo 1 Resuelto Ipc FlaxAlejandra Feliz DidierAún no hay calificaciones
- Antropologia 1er Parcial CBC UBA XxiDocumento22 páginasAntropologia 1er Parcial CBC UBA XxiSiriAún no hay calificaciones
- Biología: Guía introductoria de estudio celularDocumento44 páginasBiología: Guía introductoria de estudio celularDaniel LibedinskyAún no hay calificaciones
- El proceso de equilibración en la construcción del conocimientoDocumento14 páginasEl proceso de equilibración en la construcción del conocimientoDiana Fernandez ZalazarAún no hay calificaciones
- A. Bulcourf-Vazquez. La Ciencia Política Como ProfesiónDocumento50 páginasA. Bulcourf-Vazquez. La Ciencia Política Como ProfesiónUriel Navarro PeñalozaAún no hay calificaciones
- Climas de Argentina Condiciones Climáticas de La Republica Argentina y RelieveDocumento6 páginasClimas de Argentina Condiciones Climáticas de La Republica Argentina y Relievevictoria_aguilera_11Aún no hay calificaciones
- Argumentos y estructuraDocumento193 páginasArgumentos y estructuraIsrael SalesAún no hay calificaciones
- La Psicología y La Teoría PsicoanalíticaDocumento15 páginasLa Psicología y La Teoría PsicoanalíticaAdriana MurilloAún no hay calificaciones
- La Revolución CopernicanaDocumento15 páginasLa Revolución CopernicanaRíos JavierAún no hay calificaciones
- Taller Análisis e InferenciaDocumento4 páginasTaller Análisis e InferenciapaolisAún no hay calificaciones
- Resumen Sociologia Intensivo de Verano 2023Documento30 páginasResumen Sociologia Intensivo de Verano 2023Nicolas TorresAún no hay calificaciones
- Relaciones Entre Simios y Humanos, Diferencias y SimilitudesDocumento19 páginasRelaciones Entre Simios y Humanos, Diferencias y SimilitudesStephanie Sepúlveda0% (2)
- Comunicación 3 - Segundo Parcial UNLZDocumento41 páginasComunicación 3 - Segundo Parcial UNLZPaul BordisAún no hay calificaciones
- Desarrollo Embrionario en La Especie HumanaDocumento48 páginasDesarrollo Embrionario en La Especie HumanaJesusAún no hay calificaciones
- Resumen 1° Parcial Biología e Intro. A La Biología Celular 54Documento29 páginasResumen 1° Parcial Biología e Intro. A La Biología Celular 54Carolina ParapugnaAún no hay calificaciones
- REPASO Mol y Grupos FuncionalesDocumento15 páginasREPASO Mol y Grupos FuncionalesDaniel Eleazar Jara SifuentesAún no hay calificaciones
- Módulo 1 IPC Valente 2022 (U 1 y 2)Documento285 páginasMódulo 1 IPC Valente 2022 (U 1 y 2)Milagros Masso100% (1)
- Programación BIOFISICADocumento2 páginasProgramación BIOFISICARaúl AltamiranoAún no hay calificaciones
- Prefectura Naval Argentina: Escuela Superior de Informatica Catedra de Matematica SuperiorDocumento13 páginasPrefectura Naval Argentina: Escuela Superior de Informatica Catedra de Matematica SuperiorAlejandro Javier AcostaAún no hay calificaciones
- Psicología Una Introducción. Serie Plata. AZDocumento110 páginasPsicología Una Introducción. Serie Plata. AZKarim Karel Nasser100% (1)
- Ecosistemas - Pablo E ZapataDocumento69 páginasEcosistemas - Pablo E ZapataJose Fernando Cisneros MorenoAún no hay calificaciones
- Clase 11. Herencia Ligada Al SexoDocumento22 páginasClase 11. Herencia Ligada Al SexoEnrique DFAún no hay calificaciones
- EquinoterapiaDocumento12 páginasEquinoterapiamanuel panchoAún no hay calificaciones
- Cosmológica griega: orígenes del universoDocumento1 páginaCosmológica griega: orígenes del universoLucas Emanuel ANRIQUEZ100% (1)
- U1la Episteme de La PsicologíaDocumento17 páginasU1la Episteme de La PsicologíaJoaquin FavaAún no hay calificaciones
- Mapa Mental Del Tema Medula EspinalDocumento1 páginaMapa Mental Del Tema Medula EspinalMaría Fernanda EspinosaAún no hay calificaciones
- Seleccion Sexual e IntrasexualDocumento14 páginasSeleccion Sexual e IntrasexualDIONEBIOS89% (9)
- Definiciones de la Psicología según Bleger y CassirerDocumento90 páginasDefiniciones de la Psicología según Bleger y CassirermartinaAún no hay calificaciones
- La evolución humana: una historia en construcciónDocumento24 páginasLa evolución humana: una historia en construcciónBrayan CastilloAún no hay calificaciones
- Pedagogia MODULO I UnLzDocumento23 páginasPedagogia MODULO I UnLzEmilse van der Linder100% (1)
- Qué Es LumbriculturaDocumento22 páginasQué Es LumbriculturaMonica Del Cisne Banegas VincesAún no hay calificaciones
- Función Química Sales PDFDocumento3 páginasFunción Química Sales PDFSofia PelaezAún no hay calificaciones
- Resumen EconomiaDocumento16 páginasResumen EconomiaVikii ArcuriiAún no hay calificaciones
- Evolucion de EspeciesDocumento4 páginasEvolucion de EspeciesMarcel Antonio Vasquez Severino100% (1)
- Flujo energía ecosistemasDocumento14 páginasFlujo energía ecosistemasAlejandro Juchasara FloresAún no hay calificaciones
- Tema 5 Enzimas y VitaminasDocumento13 páginasTema 5 Enzimas y VitaminasDavid MorenoAún no hay calificaciones
- Metabolismo: Biología CelularDocumento39 páginasMetabolismo: Biología CelularKiara NascimentoAún no hay calificaciones
- Biología Celular: Metabolismo, Catabolismo, Anabolismo y EnzimasDocumento79 páginasBiología Celular: Metabolismo, Catabolismo, Anabolismo y Enzimasjulieta gataAún no hay calificaciones
- ParteaDocumento39 páginasPartealeticiaarmacedoAún no hay calificaciones
- Enzimas y Vitaminas2ºbach2018Documento9 páginasEnzimas y Vitaminas2ºbach2018PATRICIA LÓPEZ SÁNCHEZAún no hay calificaciones
- SEM I EnzimasDocumento15 páginasSEM I Enzimasapi-3837311100% (3)
- TEMA 5 Enzimas y VitaminasDocumento10 páginasTEMA 5 Enzimas y VitaminaslgarciamarcAún no hay calificaciones
- Dolor Psíquico en La Vejez.Documento39 páginasDolor Psíquico en La Vejez.Noelia KoyukAún no hay calificaciones
- Páginas Desdematerial de CatedraDocumento1 páginaPáginas Desdematerial de CatedraImpresiones CBCAún no hay calificaciones
- Hobsbawm - Capítulo 2. La Economía Cambia de RitmoDocumento12 páginasHobsbawm - Capítulo 2. La Economía Cambia de RitmoImpresiones CBCAún no hay calificaciones
- 11.partenio y Alvarez EMPRESAS RECUPERADAS EN ARGENTINA - PRODUCCIONES, ESPACIOS Y TIEMPOS DE GÉNERODocumento9 páginas11.partenio y Alvarez EMPRESAS RECUPERADAS EN ARGENTINA - PRODUCCIONES, ESPACIOS Y TIEMPOS DE GÉNEROImpresiones CBCAún no hay calificaciones
- Construyendo Una Agenda de Género en Las Políticas Públicas en Salud. Tajer, D. 2012Documento15 páginasConstruyendo Una Agenda de Género en Las Políticas Públicas en Salud. Tajer, D. 2012Maria LealAún no hay calificaciones
- 3 FoucaultDocumento4 páginas3 FoucaultImpresiones CBCAún no hay calificaciones
- 1 AlthusserDocumento8 páginas1 AlthusserImpresiones CBCAún no hay calificaciones
- Texto Programa 2024Documento20 páginasTexto Programa 2024Impresiones CBCAún no hay calificaciones
- 8.PerezRevista Historia y Justicia, 11 - 2018Documento11 páginas8.PerezRevista Historia y Justicia, 11 - 2018Impresiones CBCAún no hay calificaciones
- T.2.2.reich - El Diagnostico Psiquiatrico Como Problema EticoDocumento12 páginasT.2.2.reich - El Diagnostico Psiquiatrico Como Problema EticoImpresiones CBCAún no hay calificaciones
- 9 AchaDocumento11 páginas9 AchaImpresiones CBCAún no hay calificaciones
- Hirsch, Joachim - Globalizacion, Capital y Estado PDFDocumento117 páginasHirsch, Joachim - Globalizacion, Capital y Estado PDFAntonio Jose Mendez BurguillosAún no hay calificaciones
- D AntonioDocumento19 páginasD AntonioImpresiones CBCAún no hay calificaciones
- T.2.10.Dejours El Trabajo Entre Sufrimiento y PlacerDocumento6 páginasT.2.10.Dejours El Trabajo Entre Sufrimiento y PlacerImpresiones CBCAún no hay calificaciones
- 4.oslak REFLEXIONES SOBRE LA FORMACIN DEL ESTADO Y LA CONSTRUCCIN DE LA SOCIEDAD ARGENTINADocumento9 páginas4.oslak REFLEXIONES SOBRE LA FORMACIN DEL ESTADO Y LA CONSTRUCCIN DE LA SOCIEDAD ARGENTINAImpresiones CBCAún no hay calificaciones
- 26 Marradi - Archenti - Piovani - 2007Documento19 páginas26 Marradi - Archenti - Piovani - 2007Impresiones CBCAún no hay calificaciones
- 2.romanutti El Estado Según Foucault - Soberanía, Biopolítica y GubernamentalidadDocumento7 páginas2.romanutti El Estado Según Foucault - Soberanía, Biopolítica y GubernamentalidadImpresiones CBCAún no hay calificaciones
- Abal MedinaDocumento16 páginasAbal MedinaGustavo Signorele KehyaianAún no hay calificaciones
- 3 sANABRIADocumento7 páginas3 sANABRIAImpresiones CBCAún no hay calificaciones
- 31 Catterberg - MorenoDocumento17 páginas31 Catterberg - MorenoImpresiones CBCAún no hay calificaciones
- CLASE 09 - Unidad 5 - Primera ParteDocumento14 páginasCLASE 09 - Unidad 5 - Primera PartePato Ruiz DiazAún no hay calificaciones
- CALVEIRO - Poder y Desaparicion ResaltadoDocumento53 páginasCALVEIRO - Poder y Desaparicion ResaltadoNadia LópezAún no hay calificaciones
- Abcdefghij Fghijklm NÑO Qrstuvwxyz: AbcdefghijklmnDocumento22 páginasAbcdefghij Fghijklm NÑO Qrstuvwxyz: AbcdefghijklmnImpresiones CBCAún no hay calificaciones
- 29 Informe - Sobre - Desarrollo - Humano - Argentina - 2005. - SinopsisDocumento31 páginas29 Informe - Sobre - Desarrollo - Humano - Argentina - 2005. - SinopsisImpresiones CBCAún no hay calificaciones
- Pensar La DictaduraDocumento22 páginasPensar La DictaduraFerAún no hay calificaciones
- CLASE 10 - Unidad 5 - Segunda ParteDocumento19 páginasCLASE 10 - Unidad 5 - Segunda ParteImpresiones CBCAún no hay calificaciones
- Media y Varianza Como Variables Aleatorias PDFDocumento3 páginasMedia y Varianza Como Variables Aleatorias PDFlaurabifolcoAún no hay calificaciones
- 1.bologna Metodos Estadisticos de InvestigacionDocumento19 páginas1.bologna Metodos Estadisticos de InvestigacionImpresiones CBCAún no hay calificaciones
- 4 QuirogaDocumento7 páginas4 QuirogaImpresiones CBCAún no hay calificaciones
- Tamano y Efecto Estadistica Muñio PDFDocumento1 páginaTamano y Efecto Estadistica Muñio PDFJulietaAún no hay calificaciones
- Practica de Enlace QuimicoDocumento12 páginasPractica de Enlace Quimicocristian maycol aramburuAún no hay calificaciones
- Baja Energia 2017 PDFDocumento10 páginasBaja Energia 2017 PDFLorena BanderaAún no hay calificaciones
- Trabajo QuimicaDocumento4 páginasTrabajo QuimicaVicente AcostaAún no hay calificaciones
- Procesos de Separación Físico-MecánicosDocumento8 páginasProcesos de Separación Físico-MecánicosDaniel ZamarripaAún no hay calificaciones
- Fuerzas IntermolecularesDocumento6 páginasFuerzas IntermolecularesYuly Vanessa Caicedo PeñaAún no hay calificaciones
- Capitulo 2 Agua y PHDocumento16 páginasCapitulo 2 Agua y PHDanteManriqueAún no hay calificaciones
- Ejercicios de Reforzamiento Unidad IDocumento15 páginasEjercicios de Reforzamiento Unidad Ikatiusca rodriguezAún no hay calificaciones
- Manual de Organica - LowDocumento155 páginasManual de Organica - LowJhonatan Román Román100% (1)
- Clasificación de sustancias orgánicas e inorgánicasDocumento10 páginasClasificación de sustancias orgánicas e inorgánicasclaiutAún no hay calificaciones
- Planchas JuntasDocumento90 páginasPlanchas JuntasCid Lord ChocoAún no hay calificaciones
- Definiciones de QuimicaDocumento29 páginasDefiniciones de Quimicaal2193000754Aún no hay calificaciones
- Enlace covalente entre átomos no metálicosDocumento13 páginasEnlace covalente entre átomos no metálicosLlocclla Ccasani GloriaAún no hay calificaciones
- Universidad de Guayaquil: Facultad de Ciencias Médicas Carrera de ObstetriciaDocumento41 páginasUniversidad de Guayaquil: Facultad de Ciencias Médicas Carrera de ObstetriciaAnahi AlbanAún no hay calificaciones
- Química General Unidad 3Documento29 páginasQuímica General Unidad 3carlos andinoAún no hay calificaciones
- Fuerzas IntermolecularesDocumento29 páginasFuerzas IntermolecularesEdinson LaraAún no hay calificaciones
- PRACTICA No. 2 ENLACE QUIMICO ResultadosDocumento7 páginasPRACTICA No. 2 ENLACE QUIMICO ResultadosSaul LópezAún no hay calificaciones
- Vectorial 5Documento8 páginasVectorial 5Nicolas ArboledaAún no hay calificaciones
- Guía # 3 - Química & FísicaDocumento4 páginasGuía # 3 - Química & FísicaeliascasellesAún no hay calificaciones
- Farmacotecnia Capitulo 44Documento52 páginasFarmacotecnia Capitulo 44Paola Andrea OrtizAún no hay calificaciones
- Resumen de Fisicoquimica 20Documento13 páginasResumen de Fisicoquimica 20jennychoqueureyAún no hay calificaciones
- Química Anual Ade 2015Documento82 páginasQuímica Anual Ade 2015Jose Luis Roca Cordova100% (1)
- Química y sus ramasDocumento70 páginasQuímica y sus ramasjoanzinhoa0% (1)
- Química del enlace químicoDocumento9 páginasQuímica del enlace químicoEdwis Castañeda VelásquezAún no hay calificaciones
- Fluidos MagneticosDocumento14 páginasFluidos MagneticosEmmanuel Flores VargasAún no hay calificaciones
- Química de PolímerosDocumento148 páginasQuímica de PolímerosJhonattás Muniz de Souza0% (1)
- Cuadernillo (Olimpiada) QuimicaDocumento35 páginasCuadernillo (Olimpiada) QuimicaJuliaAún no hay calificaciones
- Taller de QuimicaDocumento3 páginasTaller de QuimicaCarolinaAún no hay calificaciones
- Sinteisis Regla Del Octeto, Estructura de Lewis y Enlaces Químicos 7°Documento10 páginasSinteisis Regla Del Octeto, Estructura de Lewis y Enlaces Químicos 7°Quimica SebastianaAún no hay calificaciones
- Temas Selectos de Fisica..Documento286 páginasTemas Selectos de Fisica..Garcia ManuelAún no hay calificaciones
- Antecedentes Práctica 2 Fuerzas Intermoleculares y Solubilidad PDFDocumento5 páginasAntecedentes Práctica 2 Fuerzas Intermoleculares y Solubilidad PDFKazuyaUedaTatsuyaAún no hay calificaciones
- Disciplina Mental: Técnicas infalibles para lograr todo lo que te propones y eliminar la pereza y la procrastinación de tu vida para siempreDe EverandDisciplina Mental: Técnicas infalibles para lograr todo lo que te propones y eliminar la pereza y la procrastinación de tu vida para siempreCalificación: 5 de 5 estrellas5/5 (3)
- La revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaDe EverandLa revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaCalificación: 5 de 5 estrellas5/5 (200)
- La metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceDe EverandLa metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceCalificación: 5 de 5 estrellas5/5 (8)
- Reconstrucción de dientes endodonciados: Pautas de actuación clínicaDe EverandReconstrucción de dientes endodonciados: Pautas de actuación clínicaCalificación: 5 de 5 estrellas5/5 (4)
- Zensorialmente : Dejá que tu cuerpo sea tu cerebroDe EverandZensorialmente : Dejá que tu cuerpo sea tu cerebroAún no hay calificaciones
- Anatomía & 100 estiramientos Esenciales (Color): Técnicas, beneficios, precauciones, consejos, tablas de series, dolenciasDe EverandAnatomía & 100 estiramientos Esenciales (Color): Técnicas, beneficios, precauciones, consejos, tablas de series, dolenciasCalificación: 4.5 de 5 estrellas4.5/5 (21)
- Fisiopatología de las enfermedades cardiovascularesDe EverandFisiopatología de las enfermedades cardiovascularesCalificación: 5 de 5 estrellas5/5 (1)
- El libro conciso de la punción seca: Manual del terapeuta para las aplicaciones en los puntos gatillo miofasciales (Color)De EverandEl libro conciso de la punción seca: Manual del terapeuta para las aplicaciones en los puntos gatillo miofasciales (Color)Calificación: 3 de 5 estrellas3/5 (2)
- Interpretación del ECG: Una Guía Práctica e Intuitiva para Aprender a Leer el ECG y Diagnosticar y Tratar ArritmiasDe EverandInterpretación del ECG: Una Guía Práctica e Intuitiva para Aprender a Leer el ECG y Diagnosticar y Tratar ArritmiasAún no hay calificaciones
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- Desastres y emergencias. Prevención, mitigación y preparaciónDe EverandDesastres y emergencias. Prevención, mitigación y preparaciónCalificación: 4 de 5 estrellas4/5 (5)
- Prescripción de ejercico físico para la saludDe EverandPrescripción de ejercico físico para la saludCalificación: 5 de 5 estrellas5/5 (1)
- Trucos para Leer la Mente de los Demás: Cómo Adivinar el Pensamiento de los Demás con Poco Esfuerzo. 2 Libros en 1 - Secretos de la Psicología Oscura, Cómo ser un Detector de MentirasDe EverandTrucos para Leer la Mente de los Demás: Cómo Adivinar el Pensamiento de los Demás con Poco Esfuerzo. 2 Libros en 1 - Secretos de la Psicología Oscura, Cómo ser un Detector de MentirasCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Dieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasDe EverandDieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasCalificación: 4 de 5 estrellas4/5 (9)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Neurociencia para vencer la depresión: La esprial ascendenteDe EverandNeurociencia para vencer la depresión: La esprial ascendenteCalificación: 4.5 de 5 estrellas4.5/5 (10)
- La vibración de las ondas cerebrales: Recuperar el ritmo de una vida saludable y felizDe EverandLa vibración de las ondas cerebrales: Recuperar el ritmo de una vida saludable y felizCalificación: 5 de 5 estrellas5/5 (7)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- Notas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesDe EverandNotas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesAún no hay calificaciones
- El péndulo de sanación: Péndulo hebreo. Investigación y sistematización de la técnicaDe EverandEl péndulo de sanación: Péndulo hebreo. Investigación y sistematización de la técnicaCalificación: 4.5 de 5 estrellas4.5/5 (27)