También podría gustarte
- Medicina del Trabajo. Agentes Físicos y disergonómicos.De EverandMedicina del Trabajo. Agentes Físicos y disergonómicos.Aún no hay calificaciones
- Art - Distrofias MuscularesDocumento11 páginasArt - Distrofias MuscularesNat LesAún no hay calificaciones
- Miopatias HereditariasDocumento28 páginasMiopatias HereditariasLizeth ArceAún no hay calificaciones
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Lectura 6 Enfermedades Por Expansión de Tripletes PDFDocumento9 páginasLectura 6 Enfermedades Por Expansión de Tripletes PDFDiego Enzo Chala GrazaAún no hay calificaciones
- Distribución de Receptores de IP3 en Músculo Esquelético HumanoDocumento48 páginasDistribución de Receptores de IP3 en Músculo Esquelético HumanoReinaldo Figueroa ContrerasAún no hay calificaciones
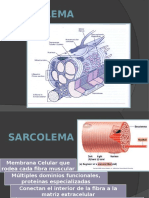
- Sarcolema: membrana muscular claveDocumento29 páginasSarcolema: membrana muscular clavetpsc20Aún no hay calificaciones
- Clasificacion_distrofiasDocumento44 páginasClasificacion_distrofiasmanu122114Aún no hay calificaciones
- Revisión: Distrofias MuscularesDocumento15 páginasRevisión: Distrofias MuscularesChristian Valera VicencioAún no hay calificaciones
- Distrofia Muscular Extremidad CinturaDocumento9 páginasDistrofia Muscular Extremidad CinturaCarlos Angel LiraAún no hay calificaciones
- Manuel Daniel Díaz MuñozDocumento174 páginasManuel Daniel Díaz MuñozElias Nassif GomezAún no hay calificaciones
- La Via Del Keap1-Nrf2Documento7 páginasLa Via Del Keap1-Nrf2Claudia RodríguezAún no hay calificaciones
- Nuevos AINEsDocumento20 páginasNuevos AINEsodontoblogger100% (3)
- 16mOP108 Reporte Completo PDFDocumento7 páginas16mOP108 Reporte Completo PDFdanicars0Aún no hay calificaciones
- Anies Mecnismo+accionDocumento4 páginasAnies Mecnismo+accionqfecrespo100% (2)
- Es Cler Oder MiaDocumento59 páginasEs Cler Oder Mialuis alfredo pernett martinezAún no hay calificaciones
- Tabla de GenesDocumento5 páginasTabla de GenesMaría José ÁlvarezAún no hay calificaciones
- Resumen Bioquimica DMDDocumento2 páginasResumen Bioquimica DMDMaria AriasAún no hay calificaciones
- Resumen Bioquimica DMDDocumento2 páginasResumen Bioquimica DMDGIOVANNA BARROSAún no hay calificaciones
- Modelo Genético de Prevención en Atención TempranaDocumento72 páginasModelo Genético de Prevención en Atención TempranaLaura PGAún no hay calificaciones
- Tarea 6 BiologiaaaaaaDocumento14 páginasTarea 6 BiologiaaaaaaJoaquin AltamiranoAún no hay calificaciones
- Escleroderma: SclerodermaDocumento9 páginasEscleroderma: SclerodermaAngélicaAún no hay calificaciones
- Fagocitosis 2019-2 PDFDocumento110 páginasFagocitosis 2019-2 PDFArturo Miñon PagazaAún no hay calificaciones
- GD Neuromuscular mgdt1219Documento6 páginasGD Neuromuscular mgdt1219Armando GBAún no hay calificaciones
- Power SeminarioDocumento59 páginasPower SeminarioblutlaboratoriosAún no hay calificaciones
- T.3. - InflamaciónDocumento88 páginasT.3. - InflamaciónEdgar Manuel Romero DíazAún no hay calificaciones
- RAE Clínico 2Documento10 páginasRAE Clínico 2xdxd.i.perrinAún no hay calificaciones
- Monografia Lumbalgia 2Documento40 páginasMonografia Lumbalgia 2estef71% (7)
- Efectos de La Terapia Fotodinámicaen La Fisiología CoroideaDocumento17 páginasEfectos de La Terapia Fotodinámicaen La Fisiología CoroidealoretoAún no hay calificaciones
- Enfermedades GenómicasDocumento13 páginasEnfermedades GenómicasIvonna VargasAún no hay calificaciones
- Fisiopatología de La Esclerosis Lateral Amiotrófica: Trabajo Fin de GradoDocumento33 páginasFisiopatología de La Esclerosis Lateral Amiotrófica: Trabajo Fin de GradoCorposalud ValdiviaAún no hay calificaciones
- Ms 041 BDocumento10 páginasMs 041 BContreras NillAún no hay calificaciones
- Cox 2Documento7 páginasCox 2Mickey VargasAún no hay calificaciones
- zbc24294 en EsDocumento14 páginaszbc24294 en EsManuel Andres Hernandez HurtadoAún no hay calificaciones
- Miopatias Apuntes ProfesorDocumento33 páginasMiopatias Apuntes Profesorapi-3698281100% (1)
- TRAD - Enfermedad de PeyronieDocumento35 páginasTRAD - Enfermedad de PeyronieAlexander Joel Guzmán CórdovaAún no hay calificaciones
- 03 Herencia MitocondrialDocumento11 páginas03 Herencia MitocondrialKarent BonillaAún no hay calificaciones
- Miostatina. Paper - En.esDocumento5 páginasMiostatina. Paper - En.esSilvia RosasAún no hay calificaciones
- Medicina 59 01112Documento7 páginasMedicina 59 01112yael1991Aún no hay calificaciones
- Craneosinostosis 1 PDFDocumento12 páginasCraneosinostosis 1 PDFWilliam Gabriel Vargas DiazAún no hay calificaciones
- Dialnet NecrosisDeCabezaFemoral 8859542Documento7 páginasDialnet NecrosisDeCabezaFemoral 8859542Silvio VollmerAún no hay calificaciones
- Cronograma Farmacologia Medica 2023-1 4 ProfesDocumento8 páginasCronograma Farmacologia Medica 2023-1 4 ProfesCamila MejiaAún no hay calificaciones
- Glut TraduccionDocumento18 páginasGlut TraduccionRaulAún no hay calificaciones
- Programa de FisiologiaDocumento5 páginasPrograma de FisiologiaOscar Espinosa AlvarezAún no hay calificaciones
- ENFERMEDADDocumento5 páginasENFERMEDADLeslye SimbañaAún no hay calificaciones
- Enfermedades Geneticas Glaucoma y Intolerancia A La LactosaDocumento9 páginasEnfermedades Geneticas Glaucoma y Intolerancia A La LactosaJosé Alberto S GAún no hay calificaciones
- Reumatologia TWCRDocumento7 páginasReumatologia TWCRCassandra CabralAún no hay calificaciones
- TJVDDocumento173 páginasTJVDYolanda VásquezAún no hay calificaciones
- Distrofia Muscular de DuchenneDocumento12 páginasDistrofia Muscular de DuchenneIvonna VargasAún no hay calificaciones
- TFG LopSanA IntervencionDocumento33 páginasTFG LopSanA IntervencionAndrea ToledoAún no hay calificaciones
- Bloquea distrofia muscular con SINE KPT-350Documento4 páginasBloquea distrofia muscular con SINE KPT-350Katherine ToalomboAún no hay calificaciones
- Tratamiento de Fracturas Diafisarias de Radio y Cúbito Con Placas PDFDocumento5 páginasTratamiento de Fracturas Diafisarias de Radio y Cúbito Con Placas PDFJulio Cesar Guillen MoralesAún no hay calificaciones
- Art. Aines e Izoformas Cox 1 y Cox2Documento10 páginasArt. Aines e Izoformas Cox 1 y Cox2Ruth OyervidesAún no hay calificaciones
- Practica1 Cromosomas GM20ADocumento3 páginasPractica1 Cromosomas GM20AMario Barragan CortezAún no hay calificaciones
- Terapéutica Con Anticuerpos MonoclonalesDocumento13 páginasTerapéutica Con Anticuerpos MonoclonalesBenestar TerapiaAún no hay calificaciones
- Art Agtr1 Nefrologia 2Documento16 páginasArt Agtr1 Nefrologia 2Irene MAún no hay calificaciones
- Enfermedad de McArdleDocumento3 páginasEnfermedad de McArdlealberto100% (1)
- Informe de Práctica 11 - Biotecnologia MolecularDocumento16 páginasInforme de Práctica 11 - Biotecnologia MolecularSEBASTIAN NOLEAún no hay calificaciones
- BQ 29-Distrofias MuscularesDocumento9 páginasBQ 29-Distrofias MuscularesMaria Alejandra Campo PlataAún no hay calificaciones
- Caso Clinico Enfermedad de ParkinsonDocumento5 páginasCaso Clinico Enfermedad de ParkinsonRuth ceronAún no hay calificaciones
- Catarata CongenitaDocumento5 páginasCatarata CongenitaeduardodubonAún no hay calificaciones
- Ambliopía OftalmoDocumento14 páginasAmbliopía OftalmoSHEYLA SHARINA FUENTES PINTOAún no hay calificaciones
- 2009 Enfermedad de Meniere Desde Las Ciencias Básicas Hasta La Medicina Clínica PDFDocumento341 páginas2009 Enfermedad de Meniere Desde Las Ciencias Básicas Hasta La Medicina Clínica PDFKimberlyn Custodio100% (1)
- FactsAbout ASTIGMATISM SP 2013Documento4 páginasFactsAbout ASTIGMATISM SP 2013José Ignacio Jorquera SierraAún no hay calificaciones
- Artículo TuberculosisDocumento16 páginasArtículo Tuberculosispaola lievanoAún no hay calificaciones
- OvariosDocumento3 páginasOvariospaola lievanoAún no hay calificaciones
- 10defectos OpticosDocumento14 páginas10defectos Opticospaola lievanoAún no hay calificaciones
- Null 2Documento12 páginasNull 2paola lievanoAún no hay calificaciones
- 2004GERARDO1 176saludeinter 130 149Documento20 páginas2004GERARDO1 176saludeinter 130 149paola lievanoAún no hay calificaciones
- 107-2017!12!06-Tema 1. Delito y Falta de Lesiones...Documento13 páginas107-2017!12!06-Tema 1. Delito y Falta de Lesiones...Citlali Jaramillo FloresAún no hay calificaciones
- PIODERMADocumento5 páginasPIODERMApaola lievanoAún no hay calificaciones
- Art. 1Documento8 páginasArt. 1paola lievanoAún no hay calificaciones
- 2004GERARDO1 176saludeinter 76 93Documento18 páginas2004GERARDO1 176saludeinter 76 93paola lievanoAún no hay calificaciones
- 18.-Helianthemum GlomeratumDocumento1 página18.-Helianthemum Glomeratumpaola lievanoAún no hay calificaciones
- 18.-Helianthemum GlomeratumDocumento2 páginas18.-Helianthemum Glomeratumpaola lievanoAún no hay calificaciones
- Cánulas de VentilaciónDocumento2 páginasCánulas de Ventilaciónpaola lievanoAún no hay calificaciones
- Cánulas de VentilaciónDocumento2 páginasCánulas de Ventilaciónpaola lievanoAún no hay calificaciones
- Pioderma GangrenosoDocumento36 páginasPioderma Gangrenosopaola lievanoAún no hay calificaciones
- Tratamientos otitis media crónicaDocumento3 páginasTratamientos otitis media crónicapaola lievanoAún no hay calificaciones
- Otitis media: clasificación, causas y complicaciones de las fases aguda, subaguda y crónicaDocumento1 páginaOtitis media: clasificación, causas y complicaciones de las fases aguda, subaguda y crónicapaola lievanoAún no hay calificaciones
- El Método Hanlon para La Determinación DE PrioridadesDocumento27 páginasEl Método Hanlon para La Determinación DE PrioridadeszhadishAún no hay calificaciones
- El Método Hanlon para La Determinación DE PrioridadesDocumento27 páginasEl Método Hanlon para La Determinación DE PrioridadeszhadishAún no hay calificaciones
- Hipoacusia NeurosensorialDocumento2 páginasHipoacusia Neurosensorialpaola lievanoAún no hay calificaciones
- Conceptos generales de diseño geométrico de carreterasDocumento5 páginasConceptos generales de diseño geométrico de carreterasAivy Jinneth CutaAún no hay calificaciones
- Herramientas Del MetalurgistaDocumento4 páginasHerramientas Del MetalurgistaNahircitaFloresCuellarAún no hay calificaciones
- Mujer emprendedoraDocumento2 páginasMujer emprendedoraLarissaMartínezAún no hay calificaciones
- WooferDocumento8 páginasWooferAngelina MorAún no hay calificaciones
- Alimentos saludables para una ciudad enfermaDocumento118 páginasAlimentos saludables para una ciudad enfermaMartha Rubi Jimenez VeraAún no hay calificaciones
- Los Concheros, La Reconquista de MexicoDocumento21 páginasLos Concheros, La Reconquista de MexicoEduardo M. Garcés100% (1)
- Taller Tema La TierraDocumento4 páginasTaller Tema La TierraRodrigo Molina ArevaloAún no hay calificaciones
- Investigacion DocumentalDocumento12 páginasInvestigacion Documentalabrajam700% (2)
- Ejercicio de LocalizacionDocumento9 páginasEjercicio de LocalizacionKevin Stiven Rocha MolinaAún no hay calificaciones
- Habitats en PeligroDocumento81 páginasHabitats en Peligroguss_1288Aún no hay calificaciones
- Co 4042Documento16 páginasCo 4042profe.williamprietoAún no hay calificaciones
- 1821-1823, Bustamante, La Abispa de ChilpancingoDocumento518 páginas1821-1823, Bustamante, La Abispa de ChilpancingoChuletasCanCanAún no hay calificaciones
- Resiliencia y Adaptabilidad OrganizacionalDocumento31 páginasResiliencia y Adaptabilidad Organizacionalsulma lopezAún no hay calificaciones
- Modulo #01 HumanidDocumento18 páginasModulo #01 HumanidCesar Meza KevinAún no hay calificaciones
- Mapa Conceptual Del MAISDocumento11 páginasMapa Conceptual Del MAISDaniel Ayala MorenoAún no hay calificaciones
- Bruselas: Ruta Art Nouveau' Por Bruselas PDFDocumento7 páginasBruselas: Ruta Art Nouveau' Por Bruselas PDFBlueOneGaussAún no hay calificaciones
- Terapias conductistasDocumento32 páginasTerapias conductistaskolob1_1000Aún no hay calificaciones
- Examen Latin Selectividad Junio 2019 Enunciado PDFDocumento3 páginasExamen Latin Selectividad Junio 2019 Enunciado PDFsoletebcAún no hay calificaciones
- Codigo Ético Del Ingeniero MecánicoDocumento2 páginasCodigo Ético Del Ingeniero MecánicoMichelle Sandoval Zamudio100% (1)
- Jorge Manrique PDFDocumento10 páginasJorge Manrique PDFAnonymous TfQapEThdAún no hay calificaciones
- 950 GCDocumento188 páginas950 GCDennys Sttefany100% (2)
- Trabajo Práctico Nº1 Pulsion - FreudDocumento8 páginasTrabajo Práctico Nº1 Pulsion - Freudmaria colmanAún no hay calificaciones
- Caso Clínico TBC VihDocumento14 páginasCaso Clínico TBC VihDangello Florian MuñozAún no hay calificaciones
- Macetas de ConcretoDocumento14 páginasMacetas de ConcretoRita Marlene50% (2)
- Evaluación CríticaDocumento9 páginasEvaluación CríticaHermes Medina50% (2)
- Plantillas AgronegociosDocumento106 páginasPlantillas AgronegociosmariopunoAún no hay calificaciones
- Tabla de La AlcoholomaníaDocumento18 páginasTabla de La AlcoholomaníaRecursos Humanos Vallercia100% (1)
- Proyecto Steam Museo LiterarioDocumento8 páginasProyecto Steam Museo LiterarioAlejandro LulorAún no hay calificaciones
- Acuifero GuaraniDocumento30 páginasAcuifero GuaraniMateo RicciardiAún no hay calificaciones
- Preguntas Frecuentes Renta de Quinta CategoriaDocumento6 páginasPreguntas Frecuentes Renta de Quinta CategoriaAura BlueAún no hay calificaciones
- Minerales y vitaminas: Micronutrientes esenciales en la alimentación, nutrición y salud.De EverandMinerales y vitaminas: Micronutrientes esenciales en la alimentación, nutrición y salud.Calificación: 3 de 5 estrellas3/5 (2)
- La química al alcance de todosDe EverandLa química al alcance de todosCalificación: 1 de 5 estrellas1/5 (1)
- Genética general: Libro de textoDe EverandGenética general: Libro de textoCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Notas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesDe EverandNotas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesAún no hay calificaciones
- Investigación de mecanismos de reacción en química orgánicaDe EverandInvestigación de mecanismos de reacción en química orgánicaAún no hay calificaciones
- Principios básicos de bioquímica de los alimentosDe EverandPrincipios básicos de bioquímica de los alimentosCalificación: 4.5 de 5 estrellas4.5/5 (2)
- Técnicas y medidas básicas en el laboratorio de químicaDe EverandTécnicas y medidas básicas en el laboratorio de químicaCalificación: 4.5 de 5 estrellas4.5/5 (6)
- Bioquímica: metabolismo energético, conceptos y aplicaciónDe EverandBioquímica: metabolismo energético, conceptos y aplicaciónCalificación: 4 de 5 estrellas4/5 (7)
- Fundamentos de química aplicados a las ciencias de la saludDe EverandFundamentos de química aplicados a las ciencias de la saludAún no hay calificaciones
- Mediciones y métodos de uso común en el laboratorio de QuímicaDe EverandMediciones y métodos de uso común en el laboratorio de QuímicaCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Lecciones de Química HeterocíclicaDe EverandLecciones de Química HeterocíclicaCalificación: 5 de 5 estrellas5/5 (1)
- Guías de laboratorio de bioquímica para la carrera de químicaDe EverandGuías de laboratorio de bioquímica para la carrera de químicaCalificación: 5 de 5 estrellas5/5 (1)
- Química orgánica: ejercicios de aplicaciónDe EverandQuímica orgánica: ejercicios de aplicaciónCalificación: 5 de 5 estrellas5/5 (3)
- Polímeros: Generalidades y tendencias de investigación en ColombiaDe EverandPolímeros: Generalidades y tendencias de investigación en ColombiaCalificación: 5 de 5 estrellas5/5 (14)
- Nomenclatura de las sustancias químicasDe EverandNomenclatura de las sustancias químicasCalificación: 5 de 5 estrellas5/5 (2)
- Estructura atómica y enlace químicoDe EverandEstructura atómica y enlace químicoCalificación: 3 de 5 estrellas3/5 (2)
- Inteligencia física: Aprende a ver el mundo con la mente de un físicoDe EverandInteligencia física: Aprende a ver el mundo con la mente de un físicoCalificación: 5 de 5 estrellas5/5 (41)
- Epigenoma: para cuidar tu cuerpo y tu vidaDe EverandEpigenoma: para cuidar tu cuerpo y tu vidaCalificación: 4 de 5 estrellas4/5 (5)
- Sistema de mejora continua de la calidad en el laboratorio: Teoría y prácticaDe EverandSistema de mejora continua de la calidad en el laboratorio: Teoría y prácticaAún no hay calificaciones
- Análisis inorgánico cualitativo sistemáticoDe EverandAnálisis inorgánico cualitativo sistemáticoCalificación: 4.5 de 5 estrellas4.5/5 (2)