También podría gustarte
- Memoria TFG. Sergio Palomino Gálvez.Documento31 páginasMemoria TFG. Sergio Palomino Gálvez.Sergio PalAún no hay calificaciones
- Genes Homeoticos en PlantasDocumento28 páginasGenes Homeoticos en PlantasMichelle Chung Blas100% (1)
- Splicing Alternativo y La Progresión TumoralDocumento17 páginasSplicing Alternativo y La Progresión TumoralUSMP FN ARCHIVOSAún no hay calificaciones
- Gametogenesis A Journey From Inception To Conception - En.esDocumento47 páginasGametogenesis A Journey From Inception To Conception - En.esLisdany BecerraAún no hay calificaciones
- Variación SomaclonalDocumento9 páginasVariación SomaclonalWillSmith SanchezAún no hay calificaciones
- Revision Biblografica GeneticaDocumento6 páginasRevision Biblografica GeneticaLAURA VANESSA MEZA ARRIETAAún no hay calificaciones
- Interacciones MAPK-tumoresDocumento5 páginasInteracciones MAPK-tumoresJesus GomezAún no hay calificaciones
- Grupo 2. Preproyecto Forked-Sepia Corregido. Carvajal, Díaz, HernándezDocumento15 páginasGrupo 2. Preproyecto Forked-Sepia Corregido. Carvajal, Díaz, HernándezDiego GarcíaAún no hay calificaciones
- Genes Homeoticos en Plantas 3Documento27 páginasGenes Homeoticos en Plantas 3Oswaldo Manuel Ramirez MarinAún no hay calificaciones
- Impronta GenómicaDocumento4 páginasImpronta GenómicaTefy Paho AyalaAún no hay calificaciones
- GeneticaDocumento9 páginasGeneticaAngela DávilaAún no hay calificaciones
- Ensayo Genetica MolecularDocumento4 páginasEnsayo Genetica MolecularNaren Mauricio TellezAún no hay calificaciones
- Clase 4 - Reproduccion Animal - Gamtogensis - FecundacionDocumento9 páginasClase 4 - Reproduccion Animal - Gamtogensis - FecundacionEvelynNicoleCutipaCalcinaAún no hay calificaciones
- Centro Genetico Del DesarrolloDocumento7 páginasCentro Genetico Del DesarrolloJazmin Adrianaa AlvarezAún no hay calificaciones
- Resumen embriologíaDocumento4 páginasResumen embriologíaYonalí MármolAún no hay calificaciones
- La Estabilidad Genetica en La Micropropagación de PlantasDocumento10 páginasLa Estabilidad Genetica en La Micropropagación de PlantasCharly BrownAún no hay calificaciones
- Variabilidad EpigenéticaDocumento12 páginasVariabilidad EpigenéticasuperdaunAún no hay calificaciones
- Diferenciación CelularDocumento14 páginasDiferenciación Celularjuanperez19tkAún no hay calificaciones
- El Establecimiento de Los Ejes CorporalesDocumento4 páginasEl Establecimiento de Los Ejes CorporalesAlfredo Jimenez0% (1)
- Apunte GeneticaDocumento39 páginasApunte GeneticaVivi GilAún no hay calificaciones
- Drosophila MelanogasterDocumento4 páginasDrosophila MelanogasterValentina ArdilaAún no hay calificaciones
- Webcn, CN66-062Documento22 páginasWebcn, CN66-062sheyla burgosAún no hay calificaciones
- Aplicación de Los Marcadores Moleculares DNA para El Mapeo CromosómicoDocumento5 páginasAplicación de Los Marcadores Moleculares DNA para El Mapeo CromosómicoMelvin L. RuizAún no hay calificaciones
- Traduccion ReloadedDocumento20 páginasTraduccion ReloadedLuisSarmientoAún no hay calificaciones
- Desarrollo Embrionario Y Diferenciacion Sexual: 1.1. SexoDocumento19 páginasDesarrollo Embrionario Y Diferenciacion Sexual: 1.1. SexojuanAún no hay calificaciones
- Ensayo MicrosatelitesDocumento3 páginasEnsayo MicrosatelitesNorman Valdéz ZamoranoAún no hay calificaciones
- Para Qué Sirven Los IntronesDocumento2 páginasPara Qué Sirven Los IntronesFranciscoFrankensteinAvelarAún no hay calificaciones
- Genetic A Bacterian ADocumento22 páginasGenetic A Bacterian ADieiKonCaiCedAún no hay calificaciones
- Marcadores MolecularesDocumento12 páginasMarcadores MolecularesGeral93Aún no hay calificaciones
- Carbon PapaDocumento9 páginasCarbon PapananaAún no hay calificaciones
- Determinación de Sexo en ReptilesDocumento3 páginasDeterminación de Sexo en ReptilesLuis CardonaAún no hay calificaciones
- Resumen Articulos de GeneticaDocumento9 páginasResumen Articulos de GeneticacarolinaAún no hay calificaciones
- Variacion SomaclonalDocumento9 páginasVariacion SomaclonalSilvia YazarethAún no hay calificaciones
- Analisis de La Lectura Del ArtículoDocumento9 páginasAnalisis de La Lectura Del ArtículoJohaNa VdAún no hay calificaciones
- Marcadores Moleculares PDFDocumento8 páginasMarcadores Moleculares PDFCindy PajoyAún no hay calificaciones
- Histologia Grupo 2Documento15 páginasHistologia Grupo 2Mirely BustamanteAún no hay calificaciones
- (Chalmel y Rolland., 2015) Vinculación de La Transcriptómica y La Proteómica en La Espermatogénesis (Nuevo)Documento9 páginas(Chalmel y Rolland., 2015) Vinculación de La Transcriptómica y La Proteómica en La Espermatogénesis (Nuevo)David Choque HuamanAún no hay calificaciones
- TransposonesDocumento4 páginasTransposonesJessica CruzAún no hay calificaciones
- Parte VDocumento192 páginasParte VManuel MarcelinoAún no hay calificaciones
- 3''. Homo y Heterotalismo (Cifientes 22-09-08)Documento8 páginas3''. Homo y Heterotalismo (Cifientes 22-09-08)Nico Ardiles GálvezAún no hay calificaciones
- Articulo Científico Sobre La Muerte Embrionaria TempranaDocumento12 páginasArticulo Científico Sobre La Muerte Embrionaria TempranaAbby KimAún no hay calificaciones
- Defectos Congénitos y MorfogénesisDocumento17 páginasDefectos Congénitos y MorfogénesisElizabethAlegria0% (1)
- El Desarrollo Embrionario y La Diferencia CelularDocumento30 páginasEl Desarrollo Embrionario y La Diferencia CelularXimena Simbaña75% (4)
- Islas de Patogenicidad TraduccionDocumento19 páginasIslas de Patogenicidad Traduccionakuma3621Aún no hay calificaciones
- Esporulacion CienciasDocumento6 páginasEsporulacion CienciasWeisner CiroAún no hay calificaciones
- Muerte Celular ProgramadaDocumento7 páginasMuerte Celular ProgramadaGabrielCamarenaAún no hay calificaciones
- Polimorfismos en proteínasDocumento4 páginasPolimorfismos en proteínasCarlos MartínezAún no hay calificaciones
- MicrosateliteDocumento7 páginasMicrosateliteRicardo SotalìnAún no hay calificaciones
- Células Germinales EmbrionariasDocumento3 páginasCélulas Germinales EmbrionariasbiogenroAún no hay calificaciones
- Islas de Patogenicidad TraduccionDocumento17 páginasIslas de Patogenicidad TraduccionCindy GarciaAún no hay calificaciones
- Degradación de La Mitocondria Paterna Después de La FecundaciónDocumento8 páginasDegradación de La Mitocondria Paterna Después de La FecundaciónBanda Santa Ceclia Banda Santa CecliaAún no hay calificaciones
- MONOGRAFIADocumento16 páginasMONOGRAFIAisavozapataAún no hay calificaciones
- Genetica Del DesarrolloDocumento3 páginasGenetica Del Desarrolloangie_kadyys6702100% (1)
- Dos Vías SensorialesDocumento5 páginasDos Vías SensorialesJessica CruzAún no hay calificaciones
- EVD 2 Bioinfo NicoleDocumento23 páginasEVD 2 Bioinfo NicoleNicole PortilloAún no hay calificaciones
- Capitulo 9 PDFDocumento38 páginasCapitulo 9 PDFMariana Figueroa AndradeAún no hay calificaciones
- Defectos congénitos y morfogénesisDocumento17 páginasDefectos congénitos y morfogénesisgiovanna ramosAún no hay calificaciones
- Más allá del ADN: La Revolución Epigenética: Desde Mecanismos Celulares hasta Factores Ambientales: Cómo la Epigenética Moldea Nuestro Destino Biológico y las Implicaciones para la Salud, el Comportamiento y el Futuro de la InvestigaciónDe EverandMás allá del ADN: La Revolución Epigenética: Desde Mecanismos Celulares hasta Factores Ambientales: Cómo la Epigenética Moldea Nuestro Destino Biológico y las Implicaciones para la Salud, el Comportamiento y el Futuro de la InvestigaciónAún no hay calificaciones
- YolandaDocumento7 páginasYolandaPineda Lazaro AlexandraAún no hay calificaciones
- Razas Comerciales de CuyesDocumento24 páginasRazas Comerciales de CuyesPineda Lazaro AlexandraAún no hay calificaciones
- El Efecto Invernadero y Lluvia ÁcidaDocumento14 páginasEl Efecto Invernadero y Lluvia ÁcidaPineda Lazaro AlexandraAún no hay calificaciones
- Deontologia 3Documento4 páginasDeontologia 3Pineda Lazaro AlexandraAún no hay calificaciones
- Tradados de Libre ComercioDocumento9 páginasTradados de Libre ComercioPineda Lazaro AlexandraAún no hay calificaciones
- Carnot CiclonDocumento26 páginasCarnot CiclonPineda Lazaro AlexandraAún no hay calificaciones
- CitrarDocumento7 páginasCitrarPineda Lazaro AlexandraAún no hay calificaciones
- Inmunología Contra El CancerDocumento12 páginasInmunología Contra El CancerPineda Lazaro AlexandraAún no hay calificaciones
- PMA-EIRD-exposición de Realidad Nacional e InternacionalDocumento17 páginasPMA-EIRD-exposición de Realidad Nacional e InternacionalPineda Lazaro AlexandraAún no hay calificaciones
- Aplicaciones biotecnológicas de la enzima invertasaDocumento2 páginasAplicaciones biotecnológicas de la enzima invertasaPineda Lazaro AlexandraAún no hay calificaciones
- Biotecnologia GrisDocumento4 páginasBiotecnologia GrisPineda Lazaro Alexandra100% (1)
- PROTEOMA DEL Homo Sapiens SapiensDocumento3 páginasPROTEOMA DEL Homo Sapiens SapiensPineda Lazaro AlexandraAún no hay calificaciones
- ECOLOGIADocumento25 páginasECOLOGIAPineda Lazaro AlexandraAún no hay calificaciones
- Tarea de Biotecnologia AnimalDocumento12 páginasTarea de Biotecnologia AnimalPineda Lazaro AlexandraAún no hay calificaciones
- Tema ChampiñonDocumento22 páginasTema ChampiñonPineda Lazaro AlexandraAún no hay calificaciones
- Cómo funciona el operón araBAD que regula la vía metabólica de las pentosas en E. coliDocumento7 páginasCómo funciona el operón araBAD que regula la vía metabólica de las pentosas en E. coliPineda Lazaro AlexandraAún no hay calificaciones
- Ascariasis: Generalidades y Ciclo de VidaDocumento24 páginasAscariasis: Generalidades y Ciclo de VidaPineda Lazaro AlexandraAún no hay calificaciones
- La Ludopatía Es Un Problema Que Azota A La Población Afectando A Nuestra SociedadDocumento1 páginaLa Ludopatía Es Un Problema Que Azota A La Población Afectando A Nuestra SociedadPineda Lazaro AlexandraAún no hay calificaciones
- Todo Tiene Un Lado PositivoDocumento1 páginaTodo Tiene Un Lado PositivoPineda Lazaro AlexandraAún no hay calificaciones
- Biotecnologia de Proteinas Recombinates para La Aplicacion de La AcuaculturaDocumento23 páginasBiotecnologia de Proteinas Recombinates para La Aplicacion de La AcuaculturaGerman Maza0% (1)
- Trabajo de Compu Ale PinedaDocumento7 páginasTrabajo de Compu Ale PinedaPineda Lazaro AlexandraAún no hay calificaciones
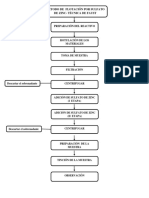
- Flujograma de Practica 5Documento1 páginaFlujograma de Practica 5Pineda Lazaro AlexandraAún no hay calificaciones
- BiotecnologiaDocumento83 páginasBiotecnologiaPineda Lazaro AlexandraAún no hay calificaciones
- Sensibilidad Del EtanolDocumento1 páginaSensibilidad Del EtanolPineda Lazaro AlexandraAún no hay calificaciones
- Semana de Fe y Meditacion en OtuzcoDocumento2 páginasSemana de Fe y Meditacion en OtuzcoPineda Lazaro AlexandraAún no hay calificaciones
- Métodos de Análisis de BiomoléculasDocumento21 páginasMétodos de Análisis de BiomoléculasPineda Lazaro AlexandraAún no hay calificaciones
- Aplicaciones biotecnológicas de la enzima invertasaDocumento2 páginasAplicaciones biotecnológicas de la enzima invertasaPineda Lazaro AlexandraAún no hay calificaciones
- Bufo PoeppigiiDocumento5 páginasBufo PoeppigiiPineda Lazaro AlexandraAún no hay calificaciones
- Prueba de Hipótesis para Muestras DependientesDocumento6 páginasPrueba de Hipótesis para Muestras DependientesPineda Lazaro AlexandraAún no hay calificaciones
- Biologia Julio2015Documento8 páginasBiologia Julio2015GipsianAún no hay calificaciones
- Cuadro Comparativo (Métodos y Técnicas de Mejoramiento Genético-Unesr)Documento7 páginasCuadro Comparativo (Métodos y Técnicas de Mejoramiento Genético-Unesr)Fredrick AlvaradoAún no hay calificaciones
- Actividades de NaturalesDocumento18 páginasActividades de NaturalesLORENA FUENTESAún no hay calificaciones
- Ibero Bio FinalDocumento92 páginasIbero Bio FinaldasdaAún no hay calificaciones
- Errores metabólicos lisosomalesDocumento9 páginasErrores metabólicos lisosomalesCarlos Yhair GuerronAún no hay calificaciones
- MicrobianaDocumento6 páginasMicrobianaJose Antonio Martinez SanchezAún no hay calificaciones
- Extracción de ADN vegetal en el laboratorio de biologíaDocumento7 páginasExtracción de ADN vegetal en el laboratorio de biologíaHernández Gómez Dayana PaolaAún no hay calificaciones
- Células MadreDocumento6 páginasCélulas MadrealejanrAún no hay calificaciones
- Mutaciones: Enfermedades Relacionadas Con El Ciclo CelularDocumento46 páginasMutaciones: Enfermedades Relacionadas Con El Ciclo CelularJorge ZanabriaAún no hay calificaciones
- MalariaDocumento14 páginasMalariaGaby NicoleAún no hay calificaciones
- Mapa Conceptual de Acidos NucleicosDocumento2 páginasMapa Conceptual de Acidos NucleicosMIGUEL ANGEL HUAMAN CUSIQUISPEAún no hay calificaciones
- Sem 11 Informe Practica 11 INTRODUCCIÓN A LA BIOINFORMÁTICA 2021-I MICROBIOLOGÍADocumento12 páginasSem 11 Informe Practica 11 INTRODUCCIÓN A LA BIOINFORMÁTICA 2021-I MICROBIOLOGÍARoberto CrepesAún no hay calificaciones
- Programa de BiologíaDocumento21 páginasPrograma de BiologíaThomas RamosAún no hay calificaciones
- Beneficios biotecnología tradicional saludDocumento3 páginasBeneficios biotecnología tradicional saludJohn Callupe ChávezAún no hay calificaciones
- Tema 16Documento8 páginasTema 16Marta AlferezAún no hay calificaciones
- Heterosis PDFDocumento206 páginasHeterosis PDFFirase Filiberto Ramos SerdaAún no hay calificaciones
- 3a Recuperacion PedagogicaDocumento7 páginas3a Recuperacion PedagogicaVictor GarciaAún no hay calificaciones
- Guia Olimpiada PDFDocumento68 páginasGuia Olimpiada PDFRobert LeeAún no hay calificaciones
- Gene Therapy For Amyotrophic Lateral Sclerosis - 2012 - Neurobiology of DiseaseDocumento7 páginasGene Therapy For Amyotrophic Lateral Sclerosis - 2012 - Neurobiology of Diseasemarieta301105Aún no hay calificaciones
- Nucleoproteinas AdnDocumento31 páginasNucleoproteinas AdnMilagrosCaballeroMedinaAún no hay calificaciones
- Beringia - Yacimiento de YanaDocumento49 páginasBeringia - Yacimiento de YanaFilosofía TextosAún no hay calificaciones
- AVANCES DE LA INGENIERIA GENÉTICA Y EL CANCER Agenda 3 BiologiaDocumento1 páginaAVANCES DE LA INGENIERIA GENÉTICA Y EL CANCER Agenda 3 Biologiamalu cavasAún no hay calificaciones
- Teoría de La MULTICELULARIDAD.Documento2 páginasTeoría de La MULTICELULARIDAD.Fernando Polo100% (1)
- El Genio Que Todos Llevamos DentroDocumento35 páginasEl Genio Que Todos Llevamos DentroYurian100% (2)
- Monografia de ARN Y ADNDocumento30 páginasMonografia de ARN Y ADNSonidos Renjifo Ccopa Condori75% (4)
- Genética: Las letras del ADN y la herencia de característicasDocumento13 páginasGenética: Las letras del ADN y la herencia de característicasJubitza VarasAún no hay calificaciones
- Trabajo Final BiologiaDocumento12 páginasTrabajo Final BiologiaCM EyjedaAún no hay calificaciones
- Excepciones de Las Leyes de MendelDocumento2 páginasExcepciones de Las Leyes de MendelSecretaria Kennedy100% (1)
- Trabajos de Mendel y Los Principios de La HerenciaDocumento10 páginasTrabajos de Mendel y Los Principios de La HerenciaJuan Carloz Ist WiedergeborenAún no hay calificaciones
- Xenotrasplantes, la solución a la escasez de órganosDocumento10 páginasXenotrasplantes, la solución a la escasez de órganosClimaco Pastor Pinza CardenasAún no hay calificaciones