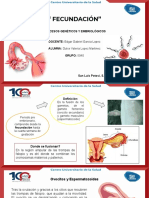
También podría gustarte
- Introducción a la Biología: RESÚMENES UNIVERSITARIOSDe EverandIntroducción a la Biología: RESÚMENES UNIVERSITARIOSCalificación: 5 de 5 estrellas5/5 (1)
- FecundacionDocumento11 páginasFecundacionDulce LopezAún no hay calificaciones
- Resumen Infectologia PDFDocumento179 páginasResumen Infectologia PDFdidáctica lengua castellana IAún no hay calificaciones
- Desarrollo EmbrionarioDocumento12 páginasDesarrollo EmbrionarioMarilyn Kilzi100% (1)
- Reproduccion Animal - 2Documento24 páginasReproduccion Animal - 2RUBEN MORALES BETANCURAún no hay calificaciones
- Ejemplos Del Método CientíficoDocumento6 páginasEjemplos Del Método CientíficoMoises Loayza Sanchez0% (1)
- GUIA DE PRACTICA N o 2 INTRODUCCION A LADocumento6 páginasGUIA DE PRACTICA N o 2 INTRODUCCION A LAMelany FranciaAún no hay calificaciones
- Genetica Del DesarrolloDocumento3 páginasGenetica Del Desarrolloangie_kadyys6702100% (1)
- Par Biomagnetico Del 109 Al 222 2a ParteDocumento21 páginasPar Biomagnetico Del 109 Al 222 2a Partejoereanimated100% (1)
- Características Generales de Moscas CorregidoDocumento27 páginasCaracterísticas Generales de Moscas CorregidoMichelle Montserrat Guzman BarronAún no hay calificaciones
- Resumen Genética Del Desarrollo 2023Documento37 páginasResumen Genética Del Desarrollo 2023Mayra Aldana Ocampo PratesAún no hay calificaciones
- Drosophila MelanogasterDocumento9 páginasDrosophila MelanogastersudroleyduAún no hay calificaciones
- Temas X y XI (Impreso)Documento26 páginasTemas X y XI (Impreso)Rodrigo SuárezAún no hay calificaciones
- Centro Genetico Del DesarrolloDocumento7 páginasCentro Genetico Del DesarrolloJazmin Adrianaa AlvarezAún no hay calificaciones
- Modelo de Desarrollo Embrionario: Drosophila MelanogasterDocumento15 páginasModelo de Desarrollo Embrionario: Drosophila MelanogasterJuan Manuel Jiménez VacasAún no hay calificaciones
- Memoria TFG. Sergio Palomino Gálvez.Documento31 páginasMemoria TFG. Sergio Palomino Gálvez.Sergio PalAún no hay calificaciones
- Drosophila MelanogasterDocumento3 páginasDrosophila MelanogasterrenezisAún no hay calificaciones
- Genes HomeoticosDocumento38 páginasGenes Homeoticosprofesorcienciaslab100% (1)
- Desarrollo Embrionario de DrosophilaDocumento14 páginasDesarrollo Embrionario de DrosophilaRicardo González DelgadoAún no hay calificaciones
- Lab 5,6Documento10 páginasLab 5,6MartecilloAún no hay calificaciones
- Procesos de gastrulación y formación de ejes en embriones de mamíferosDocumento10 páginasProcesos de gastrulación y formación de ejes en embriones de mamíferosSebastian MoranteAún no hay calificaciones
- REGULACIÓN GENÉTICA EJES CUERPODocumento4 páginasREGULACIÓN GENÉTICA EJES CUERPOAndre ChevarriaAún no hay calificaciones
- Fundamentos Biológicos Del Inicio de La Vida HumanaDocumento13 páginasFundamentos Biológicos Del Inicio de La Vida HumanaTatiana Colunche MontenegroAún no hay calificaciones
- Cuestionario EmbriologíaDocumento89 páginasCuestionario EmbriologíaDavenia SaldañaAún no hay calificaciones
- Trabajo Embriologia N°2Documento9 páginasTrabajo Embriologia N°2Diego Davila RamirezAún no hay calificaciones
- Xenopus Cap 7 ESDocumento26 páginasXenopus Cap 7 ESAlejandra GonzalezAún no hay calificaciones
- Control Genético Del DesarrolloDocumento27 páginasControl Genético Del DesarrolloEros Marin Millan100% (1)
- Resumen Organismos ModelosDocumento6 páginasResumen Organismos ModelosPaula GiraldoAún no hay calificaciones
- Consolidacion Resuelta FertilidadDocumento13 páginasConsolidacion Resuelta Fertilidadgenesis perezAún no hay calificaciones
- BiologíaDocumento31 páginasBiologíaSoledad VerasAún no hay calificaciones
- Embriologia GuiaDocumento14 páginasEmbriologia GuiaAlexis HernandezAún no hay calificaciones
- EsporulacionDocumento4 páginasEsporulacionchaperoninaAún no hay calificaciones
- GASTRULACIÓNDocumento14 páginasGASTRULACIÓNCatalina Riveros Del SolarAún no hay calificaciones
- Genética de Especificación Del Eje en Drosophila MelanogasterDocumento55 páginasGenética de Especificación Del Eje en Drosophila MelanogasterCarolina GonzalesAún no hay calificaciones
- Diferenciación Celular - Marlene BocazDocumento26 páginasDiferenciación Celular - Marlene BocazAlexis Jhair Bustamante VásquezAún no hay calificaciones
- TALLER Drosophila CIPAS BIOSTORMDocumento4 páginasTALLER Drosophila CIPAS BIOSTORMkennedyAún no hay calificaciones
- TALLER Drosophila CIPAS BIOSTORMDocumento4 páginasTALLER Drosophila CIPAS BIOSTORMkennedyAún no hay calificaciones
- GASTRULACIÓN - HistologiaDocumento10 páginasGASTRULACIÓN - HistologiaRodgerCorrêaAún no hay calificaciones
- Desarrollo EmbrionarioDocumento10 páginasDesarrollo EmbrionarioJesus Enrique Peña CoronadoAún no hay calificaciones
- Mosca de La FrutaDocumento19 páginasMosca de La FrutaLaura LunaAún no hay calificaciones
- Desarrollo EmbrionarioDocumento23 páginasDesarrollo EmbrionarioCamilo Carné Candil67% (3)
- FSH - Embriología (Clase 1 A 14)Documento282 páginasFSH - Embriología (Clase 1 A 14)Camz NagoAún no hay calificaciones
- Mecanismos Biologicos Del Desarrollo Embrionario A.DDocumento40 páginasMecanismos Biologicos Del Desarrollo Embrionario A.DJOSHUA DANIEL ARAPA BUDIEL100% (1)
- Biologia de La Concepcion Aparato Reproductor (Autoguardado)Documento16 páginasBiologia de La Concepcion Aparato Reproductor (Autoguardado)Eliana Velasco100% (1)
- Ciclo Del Epitelio Seminífero en PerrosDocumento7 páginasCiclo Del Epitelio Seminífero en PerrosMoisés Rodríguez100% (1)
- Capitulo 9. Genetica.Documento49 páginasCapitulo 9. Genetica.Mauricio GodoyAún no hay calificaciones
- Informe sobre las etapas del desarrollo embrionarioDocumento8 páginasInforme sobre las etapas del desarrollo embrionariopayalag232Aún no hay calificaciones
- Espermatozoide y Plasma SeminalDocumento24 páginasEspermatozoide y Plasma SeminalAna Marisol Pari JarroAún no hay calificaciones
- Informe Semana 3 Gy eDocumento13 páginasInforme Semana 3 Gy eLenin Gutierrez VidalAún no hay calificaciones
- Trilaminar ResumenDocumento8 páginasTrilaminar ResumenJonathan ClementeAún no hay calificaciones
- ESMEDocumento8 páginasESMEOSVIN CENTENOAún no hay calificaciones
- Diferenciación CelularDocumento21 páginasDiferenciación CelularElisa100% (5)
- Fecundación 1Documento10 páginasFecundación 1DAINNEAún no hay calificaciones
- Embriologia General Primera SemanaDocumento10 páginasEmbriologia General Primera Semanababycristi94Aún no hay calificaciones
- CicloAnfibiosDocumento7 páginasCicloAnfibiosValeria FilardiAún no hay calificaciones
- Embriologia Parte 2Documento65 páginasEmbriologia Parte 2Valerie Mendoza MendezAún no hay calificaciones
- Embrio RepasoooDocumento14 páginasEmbrio RepasoooReyes Ivan García CuevasAún no hay calificaciones
- Embriogenesis TempranaDocumento13 páginasEmbriogenesis TempranaPato RaschettiAún no hay calificaciones
- EmbriologiaDocumento8 páginasEmbriologiaEdgarGonzalezMuñozAún no hay calificaciones
- Guía de TP 2 Embrio 1 y Semanas Segmentacion e Implantacion RESUELTADocumento11 páginasGuía de TP 2 Embrio 1 y Semanas Segmentacion e Implantacion RESUELTALuz Angela MorenoAún no hay calificaciones
- Unidad 1 EmbriologiaDocumento99 páginasUnidad 1 EmbriologiaanaelisagarciaAún no hay calificaciones
- Cuestionario Celulas MadresDocumento5 páginasCuestionario Celulas MadresValerie ZimmermannAún no hay calificaciones
- Thomas Morgan y El Drosophila MelanogasterDocumento8 páginasThomas Morgan y El Drosophila MelanogasterMeytrisk GoogleAún no hay calificaciones
- Plan de Acción Caso de Clase NavarroDocumento1 páginaPlan de Acción Caso de Clase NavarroValentina ArdilaAún no hay calificaciones
- Los DMDocumento1 páginaLos DMValentina ArdilaAún no hay calificaciones
- Evidencia GA1-220501102-AA1-EV01. Cuestionario: Fundamentos Del Diseño y Teoría Del ColorDocumento2 páginasEvidencia GA1-220501102-AA1-EV01. Cuestionario: Fundamentos Del Diseño y Teoría Del ColorValentina ArdilaAún no hay calificaciones
- Syllabus Humanismo-Sociedad y EticaDocumento3 páginasSyllabus Humanismo-Sociedad y EticaValentina ArdilaAún no hay calificaciones
- Tubo D PDFDocumento1 páginaTubo D PDFValentina ArdilaAún no hay calificaciones
- Editada BG Distribuidores - Internacionales - Feb2021 AN 4Documento19 páginasEditada BG Distribuidores - Internacionales - Feb2021 AN 4Valentina ArdilaAún no hay calificaciones
- Actvidad 1aDocumento5 páginasActvidad 1aValentina ArdilaAún no hay calificaciones
- Planeación Académica - Herramientas TICDocumento1 páginaPlaneación Académica - Herramientas TICValentina ArdilaAún no hay calificaciones
- Ruta de Aprendizaje 2Documento6 páginasRuta de Aprendizaje 2Valentina ArdilaAún no hay calificaciones
- Cronograma 6561fdb5e5baa98Documento1 páginaCronograma 6561fdb5e5baa98Valentina ArdilaAún no hay calificaciones
- Herramientas TIC para La Gestión de La Seguridad 02062022Documento4 páginasHerramientas TIC para La Gestión de La Seguridad 02062022Valentina ArdilaAún no hay calificaciones
- Tubo D PDFDocumento1 páginaTubo D PDFValentina ArdilaAún no hay calificaciones
- Act. 1a Diana Beltran - ForoDocumento2 páginasAct. 1a Diana Beltran - ForoValentina ArdilaAún no hay calificaciones
- Cronograma Académico Cohorte XDocumento4 páginasCronograma Académico Cohorte XValentina ArdilaAún no hay calificaciones
- Implementación planes manejo ambiental finca experimentalDocumento9 páginasImplementación planes manejo ambiental finca experimentalValentina ArdilaAún no hay calificaciones
- FRASESDocumento8 páginasFRASESValentina ArdilaAún no hay calificaciones
- Aporte Carolina Silva DPLM Paso 3Documento3 páginasAporte Carolina Silva DPLM Paso 3Valentina ArdilaAún no hay calificaciones
- Co-Odp FinalDocumento1 páginaCo-Odp FinalValentina ArdilaAún no hay calificaciones
- Taller 2 Biol Des. I - 018Documento1 páginaTaller 2 Biol Des. I - 018Valentina ArdilaAún no hay calificaciones
- Aportes T1Documento2 páginasAportes T1Valentina ArdilaAún no hay calificaciones
- Plantilla de Trabajo - Paso 2 - Reconocer Los Procesos y Contenidos para El DPLM en La Educación InfantilDocumento8 páginasPlantilla de Trabajo - Paso 2 - Reconocer Los Procesos y Contenidos para El DPLM en La Educación InfantilValentina ArdilaAún no hay calificaciones
- Plantilla de Trabajo - Paso 1 - Reflexión DPLMDocumento4 páginasPlantilla de Trabajo - Paso 1 - Reflexión DPLMValentina ArdilaAún no hay calificaciones
- DianaC - Silva - Paso 3 - Planeación DPLMDocumento14 páginasDianaC - Silva - Paso 3 - Planeación DPLMValentina ArdilaAún no hay calificaciones
- Taller 1 de Biología Del DesarrolloDocumento2 páginasTaller 1 de Biología Del DesarrolloValentina ArdilaAún no hay calificaciones
- Practica MeiosisDocumento2 páginasPractica MeiosisValentina ArdilaAún no hay calificaciones
- Practica MeiosisDocumento3 páginasPractica MeiosisValentina ArdilaAún no hay calificaciones
- Practica MeiosisDocumento2 páginasPractica MeiosisValentina ArdilaAún no hay calificaciones
- Biol DesarrolloDocumento1 páginaBiol DesarrolloValentina ArdilaAún no hay calificaciones
- Biol DesarrolloDocumento2 páginasBiol DesarrolloValentina ArdilaAún no hay calificaciones
- División CellularDocumento16 páginasDivisión CellularValentina ArdilaAún no hay calificaciones
- Hepatitis BDocumento4 páginasHepatitis BJuan Carlos Acuña GaldamesAún no hay calificaciones
- RPM y AppDocumento97 páginasRPM y AppAnonymous JgppqCkAún no hay calificaciones
- Los Seres Vivos. Taxonomía y Nomenclatura. Los Cinco Reinos, Relaciones Evolutivas. Los Virus y Su Patología. Otras Formas AcelularesDocumento21 páginasLos Seres Vivos. Taxonomía y Nomenclatura. Los Cinco Reinos, Relaciones Evolutivas. Los Virus y Su Patología. Otras Formas Acelularesvictor gilAún no hay calificaciones
- HorticulturaDocumento32 páginasHorticulturawilliamAún no hay calificaciones
- Expo Hepatitis ArregDocumento50 páginasExpo Hepatitis ArregJennyfer CampbellAún no hay calificaciones
- Rotavirus, causante de diarrea infantilDocumento10 páginasRotavirus, causante de diarrea infantilKATHLEN RODRIGUEZ ZAMBRANOAún no hay calificaciones
- Métodos de Tinción de MicroorganismosDocumento10 páginasMétodos de Tinción de MicroorganismosMaria S Chi HernandezAún no hay calificaciones
- Genética Molecular Medicina UCM 2012-2013Documento185 páginasGenética Molecular Medicina UCM 2012-2013DelamanchaFilmsAún no hay calificaciones
- Sistema Inmune PlantasDocumento3 páginasSistema Inmune PlantasCaro Sarango Osorio100% (1)
- Preguntas Diversidad Celular y Membrana Plasmática ResueltoDocumento2 páginasPreguntas Diversidad Celular y Membrana Plasmática ResueltoEdmario De Jesus CorreiaAún no hay calificaciones
- Ibr 5Documento14 páginasIbr 5Marcial CalderonAún no hay calificaciones
- Slide MicrobiologiaDocumento17 páginasSlide MicrobiologiaLethicia NevesAún no hay calificaciones
- ASO-Latex: Ensayo para determinar Anti-estreptolisina ODocumento1 páginaASO-Latex: Ensayo para determinar Anti-estreptolisina Oede hiuamanAún no hay calificaciones
- Clavulin Suspension 250mgDocumento15 páginasClavulin Suspension 250mgFelipe HoyosAún no hay calificaciones
- Farmacología InfecciosaDocumento136 páginasFarmacología Infecciosanahomy reyesAún no hay calificaciones
- 0124 7107 Reus 20 01 00082Documento7 páginas0124 7107 Reus 20 01 00082oscar fernando leiva rojasAún no hay calificaciones
- PREGUNTAS DE EXAMENES de InmunologíaDocumento3 páginasPREGUNTAS DE EXAMENES de InmunologíaadrianaAún no hay calificaciones
- TBC y VIHDocumento2 páginasTBC y VIHBrissa Milene Baldoceda ChacaAún no hay calificaciones
- Ciliados ParameciosDocumento6 páginasCiliados ParameciosLuis Carlos BautistaAún no hay calificaciones
- Histología Del Sistema Inmune, Clase Tutor, Javier Rosas BahamondeDocumento5 páginasHistología Del Sistema Inmune, Clase Tutor, Javier Rosas BahamondeMaria JoseAún no hay calificaciones
- Escape A La Muerte CelularDocumento11 páginasEscape A La Muerte CelularJosaphat AguilarAún no hay calificaciones
- Herpes Genital OriginalDocumento16 páginasHerpes Genital OriginalVeronica FariasAún no hay calificaciones
- Inteligencia Artificial y Los HumanosDocumento1 páginaInteligencia Artificial y Los HumanosBiblioteca Carmelitas-SecundariaAún no hay calificaciones
- 4 Traduccion BacterioüfagosDocumento10 páginas4 Traduccion BacterioüfagosChuck ÜbermenschAún no hay calificaciones
- Inmunidad PAUDocumento4 páginasInmunidad PAUVirus TroyanoAún no hay calificaciones
- Mecanismos de Señalización CelularDocumento4 páginasMecanismos de Señalización CelularDafne Uribe DiazAún no hay calificaciones