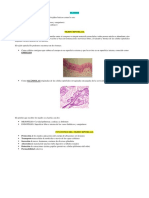
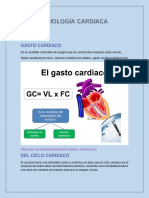
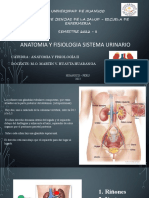
También podría gustarte
- Krebs y Glioxilato PDFDocumento6 páginasKrebs y Glioxilato PDFalexmaras9550% (4)
- Cómo Hacer La Meditación de La Sonrisa InteriorDocumento2 páginasCómo Hacer La Meditación de La Sonrisa InteriorHum MahtAún no hay calificaciones
- Hematologia PDFDocumento785 páginasHematologia PDFangel guerra100% (2)
- Desarrollo Psicológico Del NiñoDocumento75 páginasDesarrollo Psicológico Del NiñoHugo Salas L100% (1)
- Tabla de Contenidos - Urologia CIBDocumento3 páginasTabla de Contenidos - Urologia CIBolgalonAún no hay calificaciones
- Resumen de Bioquimica MetabolicaDocumento5 páginasResumen de Bioquimica MetabolicaNathalia RomeroAún no hay calificaciones
- Resumen 2 QUIMICA BIOLOGICA Original (Autoguardado)Documento11 páginasResumen 2 QUIMICA BIOLOGICA Original (Autoguardado)Inspectoria San Antonio de ArredondoAún no hay calificaciones
- Cuestionario Sistemas EnergeticosDocumento4 páginasCuestionario Sistemas EnergeticosJESUS DAVID LOPEZ PAREDESAún no hay calificaciones
- Presentación Del TemaDocumento5 páginasPresentación Del TemaRosa SanchezAún no hay calificaciones
- Ciclo de KrebsDocumento5 páginasCiclo de KrebsAndresMendozaJimenezAún no hay calificaciones
- Rutas BioquimicaDocumento6 páginasRutas BioquimicaJose lopezAún no hay calificaciones
- Metabolismo de CarbohidratosDocumento5 páginasMetabolismo de CarbohidratosMARCELA TORRICO CAMPEROSAún no hay calificaciones
- Resumen de Rutas Metabólicas BCMDocumento52 páginasResumen de Rutas Metabólicas BCMLuana HuertasAún no hay calificaciones
- (PDF) 4. - Krebs y GlioxilatoDocumento7 páginas(PDF) 4. - Krebs y GlioxilatoJean ZuñigaAún no hay calificaciones
- Metabolismo y NutriciónDocumento10 páginasMetabolismo y NutriciónMakarena BUSTAMANTEAún no hay calificaciones
- TEMA 12 Glicolisis-1Documento5 páginasTEMA 12 Glicolisis-1Majo :3Aún no hay calificaciones
- El ATP BioquimicaDocumento6 páginasEl ATP BioquimicaJose lopezAún no hay calificaciones
- Mitocondrias: órganelos clave en el metabolismo energéticoDocumento33 páginasMitocondrias: órganelos clave en el metabolismo energéticoPriscila Castillo SilvaAún no hay calificaciones
- Ciclo de KrebsDocumento2 páginasCiclo de KrebsPaola Belen LlamocaAún no hay calificaciones
- 3 Ciclo de KrebsDocumento9 páginas3 Ciclo de KrebsPaola MichaelAún no hay calificaciones
- Taller 2 2015 Bioquimica ResueltoDocumento8 páginasTaller 2 2015 Bioquimica ResueltoGisela AcostaAún no hay calificaciones
- Ciclo de Krebs y Fosforilacion OxDocumento16 páginasCiclo de Krebs y Fosforilacion Oxjoaquin pinuerAún no hay calificaciones
- Los metabolismo energéticos: FCr, glucólisis y sistema aeróbicoDocumento2 páginasLos metabolismo energéticos: FCr, glucólisis y sistema aeróbicojoseAún no hay calificaciones
- Resumen Ciclo Del Acido CitricoDocumento15 páginasResumen Ciclo Del Acido CitricoemmaAún no hay calificaciones
- Metabolismo de Carbohidratos y LipidosDocumento12 páginasMetabolismo de Carbohidratos y Lipidoscarlos100% (2)
- KrebsDocumento6 páginasKrebsAbejitas NutrilocasAún no hay calificaciones
- Cuestionario - Respiración CelularDocumento2 páginasCuestionario - Respiración CelularKateryn ÁlvarezAún no hay calificaciones
- 1.2.-Ciclo Del Ácido Cítrico - ÓsmosisDocumento4 páginas1.2.-Ciclo Del Ácido Cítrico - ÓsmosisRonald Alexander Campoverde RiosAún no hay calificaciones
- GlicolisisDocumento7 páginasGlicolisisIamChachiStephAún no hay calificaciones
- Semana 10. Ciclo de Krebs - Cte-FoDocumento52 páginasSemana 10. Ciclo de Krebs - Cte-FoVIVIANA ZULEMA AZA�ERO MAGANAún no hay calificaciones
- Glucólisis y El Ciclo de KrebsDocumento2 páginasGlucólisis y El Ciclo de KrebsFarid CastroAún no hay calificaciones
- Resumen Respiracion CelularDocumento4 páginasResumen Respiracion CelularXico EscobedoAún no hay calificaciones
- Fermentacion PDFDocumento5 páginasFermentacion PDFAnderson TorresAún no hay calificaciones
- Vías MetabólicasDocumento8 páginasVías MetabólicasKaren Justiniano Ceballos100% (1)
- Bioquímica de La NutriciónDocumento9 páginasBioquímica de La NutriciónDavid LemoineAún no hay calificaciones
- Metabolismos de Hidratos de CarbonoDocumento3 páginasMetabolismos de Hidratos de CarbonoTania Elizabeth ToledanoAún no hay calificaciones
- Ciclo de KrebsDocumento26 páginasCiclo de Krebsarmando valenciaAún no hay calificaciones
- Biología - Química UNED, Tema6Documento3 páginasBiología - Química UNED, Tema6Marta Arias FernandezAún no hay calificaciones
- Respiración CelularDocumento3 páginasRespiración CelularZaira Mía Velázquez MartínezAún no hay calificaciones
- Rutas Metabolicas, ResumenDocumento6 páginasRutas Metabolicas, ResumenLizSR01100% (2)
- Carbos BiokaDocumento13 páginasCarbos Biokaisis ortegaAún no hay calificaciones
- Vías Bioca Parcial 1Documento36 páginasVías Bioca Parcial 1isis ortegaAún no hay calificaciones
- Segundo Parcial RegMetDocumento21 páginasSegundo Parcial RegMetManuel Rueda MartinezAún no hay calificaciones
- 1 AnabolismoDocumento33 páginas1 AnabolismoWences RqtAún no hay calificaciones
- Catabolismo GlucidosDocumento3 páginasCatabolismo GlucidosAna Garcia MartínezAún no hay calificaciones
- IV-V-CARBOHIDRATOS (Autoguardado)Documento47 páginasIV-V-CARBOHIDRATOS (Autoguardado)Emelyn PeralesAún no hay calificaciones
- 3.1 Metabolismo HDCDocumento27 páginas3.1 Metabolismo HDCConstanza JaramilloAún no hay calificaciones
- Ciclo de KrebsDocumento33 páginasCiclo de Krebsdanny estelaAún no hay calificaciones
- Bioquímica 18 16 II.UDocumento29 páginasBioquímica 18 16 II.ULorena LisbethAún no hay calificaciones
- Ciclo de KrebsDocumento33 páginasCiclo de KrebsDexydeAún no hay calificaciones
- GlucolisisDocumento10 páginasGlucolisisNati SilvaAún no hay calificaciones
- TEMA 1 MetabolismoDocumento14 páginasTEMA 1 MetabolismoBerenize VidalAún no hay calificaciones
- LipidosDocumento4 páginasLipidosMarcelinho ErikAún no hay calificaciones
- Tema 8 Metabolismo Celular. BioenergéticaDocumento67 páginasTema 8 Metabolismo Celular. BioenergéticaGabriela DiazAún no hay calificaciones
- Guia MetabolismoDocumento16 páginasGuia MetabolismoVane ReyesAún no hay calificaciones
- AFPD2 Cómo Las Células Obtienen Energía de Los AlimentosDocumento21 páginasAFPD2 Cómo Las Células Obtienen Energía de Los AlimentosJIMENA ORTIZ SALDAÑAAún no hay calificaciones
- Ciclo Krebs y AtpDocumento5 páginasCiclo Krebs y AtpRONALDAún no hay calificaciones
- Gluconeogénesis: formación de glucosa a partir de precursores no glucosídicosDocumento14 páginasGluconeogénesis: formación de glucosa a partir de precursores no glucosídicosYesseniaAún no hay calificaciones
- Calculo de ATP Por Héctor LezcanoDocumento5 páginasCalculo de ATP Por Héctor LezcanoYui Dian Misa LizAún no hay calificaciones
- GlucolisisDocumento26 páginasGlucolisisAle JoyaAún no hay calificaciones
- Mecanismos de Obtención de EnergíaDocumento11 páginasMecanismos de Obtención de EnergíaBlanca CamarinhaAún no hay calificaciones
- El Ciclo de KrepsDocumento18 páginasEl Ciclo de KrepsJesus VillasanaAún no hay calificaciones
- Calculos de AtpDocumento5 páginasCalculos de AtpKaren Yohanna Cruz RodriguezAún no hay calificaciones
- Catabolismo de ProteínasDocumento14 páginasCatabolismo de ProteínasCarlos ChiaAún no hay calificaciones
- Metabolismo Del GlucógenoDocumento7 páginasMetabolismo Del GlucógenoCarlos ChiaAún no hay calificaciones
- Generalidades Del MetabolismoDocumento19 páginasGeneralidades Del MetabolismoCarlos ChiaAún no hay calificaciones
- Inhibidor del complejo I: Sodico AmitalDocumento6 páginasInhibidor del complejo I: Sodico AmitalCarlos ChiaAún no hay calificaciones
- GLÚCIDOSDocumento2 páginasGLÚCIDOSCarlos ChiaAún no hay calificaciones
- Óxido Nítrico - BiogenéticaDocumento3 páginasÓxido Nítrico - BiogenéticaCarlos ChiaAún no hay calificaciones
- Casos Practicos de Alteraciones Del Equilibrio Acido-Base PDFDocumento5 páginasCasos Practicos de Alteraciones Del Equilibrio Acido-Base PDFluzalegriaostosAún no hay calificaciones
- Taller de Membranas Biológicas PDFDocumento9 páginasTaller de Membranas Biológicas PDFCarlos ChiaAún no hay calificaciones
- Disforia de Género - GenéticaDocumento3 páginasDisforia de Género - GenéticaCarlos ChiaAún no hay calificaciones
- Disforia de Género - GenéticaDocumento3 páginasDisforia de Género - GenéticaCarlos ChiaAún no hay calificaciones
- Marcadores TumoralesDocumento7 páginasMarcadores TumoralesCarlos ChiaAún no hay calificaciones
- Generalidades Tejido EpitelialDocumento5 páginasGeneralidades Tejido EpitelialCarlos ChiaAún no hay calificaciones
- Errores de Implantación y PreeclampsiaDocumento2 páginasErrores de Implantación y PreeclampsiaCarlos ChiaAún no hay calificaciones
- ParkinsonDocumento1 páginaParkinsonCarlos ChiaAún no hay calificaciones
- Inhibidores del complejo I: RotenonaDocumento8 páginasInhibidores del complejo I: RotenonaCarlos ChiaAún no hay calificaciones
- Técnica Histológica - TincionesDocumento7 páginasTécnica Histológica - TincionesCarlos ChiaAún no hay calificaciones
- Virus y CiclosDocumento2 páginasVirus y CiclosCarlos ChiaAún no hay calificaciones
- Fibrosis QuisticaDocumento4 páginasFibrosis QuisticaCarlos ChiaAún no hay calificaciones
- SISTEMA ÓSEO (Generalidades - Huesos Del Cráneo)Documento8 páginasSISTEMA ÓSEO (Generalidades - Huesos Del Cráneo)Carlos ChiaAún no hay calificaciones
- Respiración Celular - GlosarioDocumento3 páginasRespiración Celular - GlosarioCarlos ChiaAún no hay calificaciones
- GLÚCIDOSDocumento2 páginasGLÚCIDOSCarlos ChiaAún no hay calificaciones
- Imagenes Microbiologia de La PielDocumento40 páginasImagenes Microbiologia de La PielCarlos ChiaAún no hay calificaciones
- Generalidades Tejido EpitelialDocumento5 páginasGeneralidades Tejido EpitelialCarlos ChiaAún no hay calificaciones
- RAQUITISMODocumento1 páginaRAQUITISMOCarlos ChiaAún no hay calificaciones
- Casos VirusDocumento24 páginasCasos VirusCarlos ChiaAún no hay calificaciones
- Placas de Epitelios, Tejidos, Hueso, Cartilago y MusculoDocumento126 páginasPlacas de Epitelios, Tejidos, Hueso, Cartilago y MusculoCarlos ChiaAún no hay calificaciones
- Pares craneales y sus funcionesDocumento4 páginasPares craneales y sus funcionesKike De León0% (1)
- Caso Clínico de Dolor y FiebreDocumento12 páginasCaso Clínico de Dolor y FiebreFabrizio Falcon CeferinoAún no hay calificaciones
- MTMDocumento47 páginasMTMmovarick100% (1)
- Colocación de Campos QuirurgicosDocumento7 páginasColocación de Campos QuirurgicosPaulina AjilaAún no hay calificaciones
- Fisiologia CardiacaDocumento18 páginasFisiologia Cardiacaapi-425369693Aún no hay calificaciones
- Control Motor LumbarDocumento28 páginasControl Motor LumbarnuxionAún no hay calificaciones
- Tecnicas Instrumentcion EndoDocumento9 páginasTecnicas Instrumentcion EndoznachchAún no hay calificaciones
- Jornada TerapéuticaDocumento29 páginasJornada TerapéuticaNatalia LargoAún no hay calificaciones
- Elaboración morcilla microondasDocumento32 páginasElaboración morcilla microondasJhurema B ChAún no hay calificaciones
- Nociones Básicas de Canto - Web Oficial de Anima AdversaDocumento5 páginasNociones Básicas de Canto - Web Oficial de Anima AdversaPakito Gomis AlagardaAún no hay calificaciones
- Fisiopatologia Del Sindrome ConvulsivoDocumento19 páginasFisiopatologia Del Sindrome ConvulsivoSociedad Científica de Estudiantes de Medicina de Ica100% (23)
- Sistema UrinarioDocumento17 páginasSistema UrinarioENRIQUE URETA QUIROZAún no hay calificaciones
- Chanque ACUICULTURADocumento25 páginasChanque ACUICULTURAEduardo Jorge Quesada HuaytaAún no hay calificaciones
- Músculos Del AntebrazoDocumento58 páginasMúsculos Del AntebrazoGREEK NEZARETH FARFAN OCHOAAún no hay calificaciones
- Rutinas Aeróbicas de NataciónDocumento4 páginasRutinas Aeróbicas de NataciónAlexAún no hay calificaciones
- Características anatómicas del hueso temporal con menos deDocumento9 páginasCaracterísticas anatómicas del hueso temporal con menos descarleth solanoAún no hay calificaciones
- Listeria SPPDocumento21 páginasListeria SPPSegundo FernandezAún no hay calificaciones
- Paper 3 Biometria y Constantes Clinicas Del Bovino Criollo en Puno PDFDocumento5 páginasPaper 3 Biometria y Constantes Clinicas Del Bovino Criollo en Puno PDFEstela Nelly Quispe RequenaAún no hay calificaciones
- Transfusiones Serieroja Internet)Documento146 páginasTransfusiones Serieroja Internet)GuillermoRoquésAún no hay calificaciones
- Escopolamina y sus efectosDocumento14 páginasEscopolamina y sus efectosmardel2009Aún no hay calificaciones
- Glandulas MucosasDocumento4 páginasGlandulas MucosasDONALDO RODRIGUEZ PEINADO100% (1)
- Qué Son Los Tejidos FundamentalesDocumento4 páginasQué Son Los Tejidos FundamentalesDorcas Sosa Mendoza100% (1)
- Consideraciones Generales de Los EndopostesDocumento54 páginasConsideraciones Generales de Los EndopostesLinda Rodriguez100% (1)
- Acidente de TransitoDocumento46 páginasAcidente de TransitoJuan Carlos Ayala VerduguezAún no hay calificaciones
- Mitoc. oxidación biológicaDocumento45 páginasMitoc. oxidación biológicaYadilka ViñolesAún no hay calificaciones
- Coloides, Abner Lozano LosadaDocumento13 páginasColoides, Abner Lozano Losada.....Aún no hay calificaciones