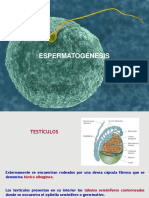
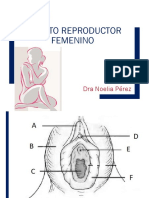
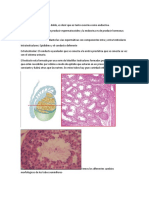
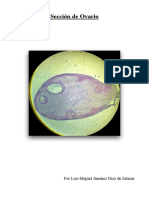
También podría gustarte
- Aparato Reproductor FemeninoDocumento77 páginasAparato Reproductor FemeninoUSMP FN ARCHIVOS100% (1)
- Embarazo, parto y lactancia del bebéDe EverandEmbarazo, parto y lactancia del bebéAún no hay calificaciones
- EMBRIOLOGIA GUIA ENURM Embriologia CompletoDocumento18 páginasEMBRIOLOGIA GUIA ENURM Embriologia CompletoMADDYAún no hay calificaciones
- Embriología GeneralDocumento23 páginasEmbriología Generalnicolas torresAún no hay calificaciones
- Gametogénesis y embriología del aparato genital femeninoDocumento12 páginasGametogénesis y embriología del aparato genital femeninoAlma Linda Peréz Santeliz100% (1)
- Anatomía e Histología Del Aparato Reproductor FemeninoDocumento74 páginasAnatomía e Histología Del Aparato Reproductor FemeninoCarlos Burgos Guerrero100% (2)
- Aparato Reproductor Femenino Trabajo ImprimirDocumento29 páginasAparato Reproductor Femenino Trabajo ImprimirLuisAún no hay calificaciones
- Histología del sistema reproductor femeninoDocumento13 páginasHistología del sistema reproductor femeninoJesmi AlemparteAún no hay calificaciones
- Aparato Reproductor FemeninoDocumento17 páginasAparato Reproductor FemeninoJuan Esteban GarciaAún no hay calificaciones
- Histología del aparato genital femenino: Ovario y úteroDocumento8 páginasHistología del aparato genital femenino: Ovario y úteroIsbelia Valentina Rivas MolinaAún no hay calificaciones
- Sistema Genital Femenino IDocumento63 páginasSistema Genital Femenino IValery ArdonAún no hay calificaciones
- Aparato Reproductor FemeninoDocumento20 páginasAparato Reproductor FemeninoKoa NunchiAún no hay calificaciones
- Fran Pregunatass VariasDocumento45 páginasFran Pregunatass VariasfkcontAún no hay calificaciones
- Sistema Genital FemeninoDocumento37 páginasSistema Genital FemeninoClaude RivasAún no hay calificaciones
- Aparato Reproductor FemeninoDocumento10 páginasAparato Reproductor FemeninoMANRIQUE GUTIÉRREZ STEPHANY GIOMAYRAAún no hay calificaciones
- Aparato Reproductor FemeninoDocumento11 páginasAparato Reproductor FemeninoChessmaster98Aún no hay calificaciones
- 9 Resumen - Síntesis - Unidad 3 - Ross-Pawlina - Histología - Capitulo 23 - Sistema Genital Femenino (1)Documento17 páginas9 Resumen - Síntesis - Unidad 3 - Ross-Pawlina - Histología - Capitulo 23 - Sistema Genital Femenino (1)Miranda LeonettiAún no hay calificaciones
- Ciclo Sexual y Ovarios. OvariosDocumento9 páginasCiclo Sexual y Ovarios. Ovariosmaria nochettiAún no hay calificaciones
- EmbriologíaDocumento17 páginasEmbriologíaCarmen Porras Jarufe50% (2)
- Aparato Reproductor FemeninoDocumento13 páginasAparato Reproductor FemeninoorlandoAún no hay calificaciones
- Aparato Reproductor FemeninoDocumento14 páginasAparato Reproductor FemeninoAngeline Cerezo Escobar100% (1)
- Procesos reproductivos masculinos: Espermatogénesis y formación de espermatozoidesDocumento53 páginasProcesos reproductivos masculinos: Espermatogénesis y formación de espermatozoidesAmanda Hernández CuadraAún no hay calificaciones
- Embrioapuntes (Parte I) - Diego MeléndezDocumento16 páginasEmbrioapuntes (Parte I) - Diego Meléndezdiego melendezAún no hay calificaciones
- Sistema Reproductor FemeninoDocumento78 páginasSistema Reproductor FemeninoIVONXYNAún no hay calificaciones
- Aparato Reproductor FemeninoDocumento25 páginasAparato Reproductor FemeninolinstilmAún no hay calificaciones
- Aparato Reproductor Femenino 2020-PARTE1 PDFDocumento51 páginasAparato Reproductor Femenino 2020-PARTE1 PDFMarcos DíasAún no hay calificaciones
- ChloeDocumento10 páginasChloeSovietWantanAún no hay calificaciones
- OVARIO HistologíaDocumento10 páginasOVARIO HistologíaAstridAún no hay calificaciones
- Ciclo Ovarico PDFDocumento6 páginasCiclo Ovarico PDFTania Alba MedranoAún no hay calificaciones
- EmbriologíaDocumento24 páginasEmbriologíaJuan Andrés RestrepoAún no hay calificaciones
- Sistema ReproductorDocumento17 páginasSistema ReproductorGianna ManziAún no hay calificaciones
- Resumen Ross-Pawlina - Histología - Capitulo 23 - Sistema Genital FemeninoDocumento17 páginasResumen Ross-Pawlina - Histología - Capitulo 23 - Sistema Genital Femeninovictoriaetchegoyen123Aún no hay calificaciones
- #10 BiologíaDocumento68 páginas#10 BiologíaMorrigan ThanatosAún no hay calificaciones
- Aparato Reproductor HembraDocumento46 páginasAparato Reproductor HembraGonzalo MunAún no hay calificaciones
- Lección No 1. Fisiología de La Reproduccion1Documento29 páginasLección No 1. Fisiología de La Reproduccion1vimarjuAún no hay calificaciones
- Trompas de Falopio y OvarioDocumento44 páginasTrompas de Falopio y OvarioAna LopezAún no hay calificaciones
- S. R. FemeninoDocumento55 páginasS. R. FemeninoErgueta Villarroel MelissaAún no hay calificaciones
- Sist. Rep. FemeninoDocumento69 páginasSist. Rep. FemeninoUSMP FN ARCHIVOSAún no hay calificaciones
- 03 Gónadas Femeninas - Origen Embriológico - Ovogénesis - Ciclos EstralesDocumento23 páginas03 Gónadas Femeninas - Origen Embriológico - Ovogénesis - Ciclos EstralesLucía María BradleyAún no hay calificaciones
- Sistema reproductor femenino: órganos y folículos ováricosDocumento28 páginasSistema reproductor femenino: órganos y folículos ováricosvalentinaAún no hay calificaciones
- Cap22Sistema GenitalDocumento30 páginasCap22Sistema GenitalPaolaAún no hay calificaciones
- Gametogénesis, Ciclo Ovarico y Ciclo SexualDocumento7 páginasGametogénesis, Ciclo Ovarico y Ciclo SexualMaia ArraizAún no hay calificaciones
- Generalidades Del Aparato Genital Femenino OvarioDocumento53 páginasGeneralidades Del Aparato Genital Femenino OvarioLuis Martinez100% (1)
- Reproductor femeninoDocumento33 páginasReproductor femeninoPAZ MARTINA ALLENDESAún no hay calificaciones
- C GinecologicaDocumento18 páginasC GinecologicaluisAún no hay calificaciones
- OVOGENESISDocumento27 páginasOVOGENESISAnonymous br1aA2Jka50% (2)
- Sistemas ReproductoresDocumento4 páginasSistemas ReproductoresntellezrAún no hay calificaciones
- Ciclo Sexual y GametogénesisDocumento5 páginasCiclo Sexual y GametogénesisSofía GilibertiAún no hay calificaciones
- 29-Aparato Reproductor FemeninoDocumento13 páginas29-Aparato Reproductor FemeninoShayAlarcónAún no hay calificaciones
- Establecimiento de La GestacionDocumento7 páginasEstablecimiento de La GestacionmarizhgAún no hay calificaciones
- Cuestionario 18Documento8 páginasCuestionario 18Eder Mendoza RoqueAún no hay calificaciones
- Capitulo 23 Sistema Genital FemeninoDocumento16 páginasCapitulo 23 Sistema Genital FemeninoJohan ChoqueAún no hay calificaciones
- UABP 2, 6 Y 9 - EmbriologíaDocumento15 páginasUABP 2, 6 Y 9 - EmbriologíaGiovanni MedranoAún no hay calificaciones
- Loe11 FisiologiaDocumento30 páginasLoe11 FisiologiaJuan Pablo Giraldo MontoyaAún no hay calificaciones
- Embriologia BasicaDocumento22 páginasEmbriologia BasicaJose AvilaAún no hay calificaciones
- Espermatogénesis y reproducción humanaDocumento33 páginasEspermatogénesis y reproducción humanaDaisy Lorena Aquino FigueredoAún no hay calificaciones
- Resumen GametogenesisDocumento7 páginasResumen GametogenesisINSTITUTO MEDERIAún no hay calificaciones
- Histología Capitulo 22Documento3 páginasHistología Capitulo 22MarceloAisaAún no hay calificaciones
- Tema 1 Desarrollo EmbrionarioDocumento19 páginasTema 1 Desarrollo EmbrionarioAnder GroundAún no hay calificaciones
- Algoritmo Clínico para El Manejo de Probable Preeclampsia Severa Menos 34 SemanasDocumento2 páginasAlgoritmo Clínico para El Manejo de Probable Preeclampsia Severa Menos 34 SemanasSteven Catunta ChoqueAún no hay calificaciones
- TUBERCULOSISDocumento35 páginasTUBERCULOSISSteven Catunta ChoqueAún no hay calificaciones
- PreeclampsiaDocumento19 páginasPreeclampsiaSteven Catunta ChoqueAún no hay calificaciones
- Placenta Previa: Factores de Riesgo y DiagnósticoDocumento14 páginasPlacenta Previa: Factores de Riesgo y DiagnósticoSteven Catunta ChoqueAún no hay calificaciones
- Manejo de PreeclampsiaDocumento1 páginaManejo de PreeclampsiaSteven Catunta ChoqueAún no hay calificaciones
- UnidoDocumento4 páginasUnidoSteven Catunta ChoqueAún no hay calificaciones
- Análisis de sangre: qué significan los resultados de una analíticaDocumento6 páginasAnálisis de sangre: qué significan los resultados de una analíticaSteven Catunta ChoqueAún no hay calificaciones
- Lesiones Invasoras Etapas 1-3Documento2 páginasLesiones Invasoras Etapas 1-3Steven Catunta ChoqueAún no hay calificaciones
- Vias de Conduccion M.EDocumento2 páginasVias de Conduccion M.ESteven Catunta ChoqueAún no hay calificaciones
- Análisis Del Caso Clínico (Tratamiento)Documento1 páginaAnálisis Del Caso Clínico (Tratamiento)Steven Catunta ChoqueAún no hay calificaciones
- Soapecha de C.a-Confirmacion de DiagnosticoDocumento1 páginaSoapecha de C.a-Confirmacion de DiagnosticoSteven Catunta ChoqueAún no hay calificaciones
- Inhibidores de La Bomba de Protones Recomendaciones Mayo 2016 PDFDocumento12 páginasInhibidores de La Bomba de Protones Recomendaciones Mayo 2016 PDFMaria Lidia Mendoza BazoaltoAún no hay calificaciones
- Patologias Frecuentes de OrganosDocumento4 páginasPatologias Frecuentes de OrganosSteven Catunta ChoqueAún no hay calificaciones
- OVIDUCTODocumento19 páginasOVIDUCTOMaritzel RiveraAún no hay calificaciones
- Balotario de Ciencia para El Examen FinalDocumento31 páginasBalotario de Ciencia para El Examen FinalDheybi Martinez RiojasAún no hay calificaciones
- Ginecologia - Conceptos BasicosDocumento3 páginasGinecologia - Conceptos BasicosRough Cibernus93% (29)
- SRM y SRFDocumento3 páginasSRM y SRFJose CelisAún no hay calificaciones
- Sistema genital femenino: anatomía y funcionesDocumento26 páginasSistema genital femenino: anatomía y funcionesWen ÁngelAún no hay calificaciones
- TP9 EsplacnoDocumento16 páginasTP9 EsplacnopabloAún no hay calificaciones
- Partes del pene y escrotoDocumento6 páginasPartes del pene y escrotoPaula EspañaAún no hay calificaciones
- SisreproDocumento52 páginasSisreproAvril AguilarAún no hay calificaciones
- Glosario EmbriologíaDocumento7 páginasGlosario EmbriologíaSalomé LuqueAún no hay calificaciones
- Clases de La Semana Del 11-05-2020 Al 15-05-2020 Docente Vinayde Duque Proyecto: Familia y Colegio Unidos Vía Digital Por Mi Educación MatemáticaDocumento3 páginasClases de La Semana Del 11-05-2020 Al 15-05-2020 Docente Vinayde Duque Proyecto: Familia y Colegio Unidos Vía Digital Por Mi Educación MatemáticaJahannay Sley Fajardo JaimesAún no hay calificaciones
- Moore - Ciclo OváricoDocumento8 páginasMoore - Ciclo OváricoFernando Junior Parra UchasaraAún no hay calificaciones
- Desarrollo Embrionario y Sus Etapas-FusionadoDocumento381 páginasDesarrollo Embrionario y Sus Etapas-FusionadoFlor Orquidea Batista SuárezAún no hay calificaciones
- Sistema circulatorio comparado en animalesDocumento56 páginasSistema circulatorio comparado en animalesBy YadhiraAún no hay calificaciones
- Sistema Reproductor Femenino v2.0Documento34 páginasSistema Reproductor Femenino v2.0Nataly Sofia Poma PerezAún no hay calificaciones
- Anatomía y fisiología del aparato reproductor femeninoDocumento13 páginasAnatomía y fisiología del aparato reproductor femeninoMaría Alejandra PeñaAún no hay calificaciones
- Guia Obste I CompletaDocumento14 páginasGuia Obste I Completaisamar duranAún no hay calificaciones
- Embriologia ResumenDocumento159 páginasEmbriologia Resumenmoleli560Aún no hay calificaciones
- Examen genital masculinoDocumento33 páginasExamen genital masculinoRandal CGAún no hay calificaciones
- Cuestionario de EmbriologiaDocumento6 páginasCuestionario de EmbriologiaEstheysi Mayerli Mamani EspinalAún no hay calificaciones
- Monografia Metodos AnticonceptivosDocumento58 páginasMonografia Metodos AnticonceptivosJonh Alexander Flores ParedesAún no hay calificaciones
- Practico 3 de BiologiaDocumento8 páginasPractico 3 de BiologiaEzequiel ZanolliAún no hay calificaciones
- Anatomia Del UteroDocumento3 páginasAnatomia Del Uteroisamar ariasAún no hay calificaciones
- Duración de la gestación en especies animalesDocumento25 páginasDuración de la gestación en especies animalesJaz SunAún no hay calificaciones
- GMB Exudado + CardíacoDocumento20 páginasGMB Exudado + Cardíacolorenaarcas23Aún no hay calificaciones
- 24 Sistema ReproductorDocumento50 páginas24 Sistema Reproductoradrian galarzaAún no hay calificaciones
- Anatomía del aparato reproductor macho y hembraDocumento33 páginasAnatomía del aparato reproductor macho y hembraYURANI VAZQUEZ AGUILERAAún no hay calificaciones
- Organos genitales internos y externos femeninosDocumento6 páginasOrganos genitales internos y externos femeninosNataly IzaAún no hay calificaciones
- Aparato Reproductor FemeninoDocumento4 páginasAparato Reproductor FemeninoAlanis Ximena EgertAún no hay calificaciones
- Sexualidad y autocuidadoDocumento72 páginasSexualidad y autocuidadoRONALD MASAVEUAún no hay calificaciones