También podría gustarte
- Histología y Embrología del Sitema NerviosoDe EverandHistología y Embrología del Sitema NerviosoCalificación: 4 de 5 estrellas4/5 (4)
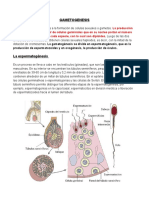
- Gametogenesis M1.3Documento10 páginasGametogenesis M1.3Hector Efrain Garcia ChavezAún no hay calificaciones
- Qué Es La GametogénesisDocumento3 páginasQué Es La GametogénesisRubenAún no hay calificaciones
- Resumen Sadler, T.W - Embriología - Capitulo 2Documento10 páginasResumen Sadler, T.W - Embriología - Capitulo 2victoriaetchegoyen123Aún no hay calificaciones
- 1 Resumen de GametogenesisDocumento8 páginas1 Resumen de GametogenesiscamiialarcondelfanteAún no hay calificaciones
- Sistema ResumidoDocumento24 páginasSistema ResumidoJorge RamirezAún no hay calificaciones
- Nociones EmbriologicasDocumento4 páginasNociones EmbriologicassarahyAún no hay calificaciones
- Proceso GametogénesisDocumento4 páginasProceso GametogénesisAlexander EztradaAún no hay calificaciones
- EmbriologíaDocumento24 páginasEmbriologíaDaniAún no hay calificaciones
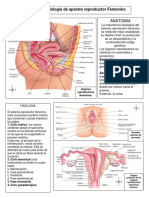
- Gametogénesis y embriología del aparato genital femeninoDocumento12 páginasGametogénesis y embriología del aparato genital femeninoAlma Linda Peréz Santeliz100% (1)
- Embrioapuntes (Parte I) - Diego MeléndezDocumento16 páginasEmbrioapuntes (Parte I) - Diego Meléndezdiego melendezAún no hay calificaciones
- Gametogénesis I y IIDocumento19 páginasGametogénesis I y IINotnow MaybelaterAún no hay calificaciones
- Células Germinales PrimordialesDocumento11 páginasCélulas Germinales Primordialesmariferc319Aún no hay calificaciones
- Fisiología de GametogénesisDocumento5 páginasFisiología de Gametogénesispier manuel cabezas armellonAún no hay calificaciones
- Capitulo 2 - Langman - GametogénesisDocumento5 páginasCapitulo 2 - Langman - Gametogénesismaria jose sanchez gilAún no hay calificaciones
- El Pibe Resumen de FecundaciónDocumento29 páginasEl Pibe Resumen de FecundaciónAntonella Violo100% (1)
- GAMETOGENÉSISDocumento6 páginasGAMETOGENÉSISanon_757572911Aún no hay calificaciones
- Espermatogénesis y OvogenesisDocumento25 páginasEspermatogénesis y OvogenesisIsrael Silva SaucedoAún no hay calificaciones
- Gametogénesis: Formación de gametos masculinos y femeninosDocumento3 páginasGametogénesis: Formación de gametos masculinos y femeninosErik Lopez MamaniAún no hay calificaciones
- Resumen Final Embriología.Documento17 páginasResumen Final Embriología.Nazareth PacayutAún no hay calificaciones
- Resumen Embriologia LangmanDocumento13 páginasResumen Embriologia Langmanmanuel_aguirre_13Aún no hay calificaciones
- Fundamentos Dela Reproducción Humana.Documento10 páginasFundamentos Dela Reproducción Humana.Iris MendozaAún no hay calificaciones
- EmbrioDocumento231 páginasEmbrioOmar C. RodríguezAún no hay calificaciones
- Apuntes embriologíaDocumento9 páginasApuntes embriologíasebastianAún no hay calificaciones
- Guía 2do Parcial de Histología, Embriología y GenéticaDocumento17 páginasGuía 2do Parcial de Histología, Embriología y GenéticaKim ZoldyckAún no hay calificaciones
- EmbriologíaDocumento17 páginasEmbriologíaCarmen Porras Jarufe50% (2)
- Resumo Embriologia MADocumento20 páginasResumo Embriologia MAabdelbrasileiro2001Aún no hay calificaciones
- GametogenesisDocumento10 páginasGametogenesisSigmaSAún no hay calificaciones
- Guía Embriología 1er ParcialDocumento35 páginasGuía Embriología 1er ParcialAlberto FongAún no hay calificaciones
- Cuadro Comparativo Entre Ovogenesis y EspermatogenesisDocumento5 páginasCuadro Comparativo Entre Ovogenesis y Espermatogenesisgavio_ta26100% (1)
- Copia de Minimalist Indian Palette Aesthetic Agency by SlidesgoDocumento15 páginasCopia de Minimalist Indian Palette Aesthetic Agency by SlidesgoGONZALEZ BERNAL EDITH MONSERRATHAún no hay calificaciones
- EmbriologíaDocumento230 páginasEmbriologíaMelissa PomaAún no hay calificaciones
- Gametogenesis OvogenesisDocumento33 páginasGametogenesis OvogenesisFernanda SotoAún no hay calificaciones
- Gametogénesis y ciclo sexual.docxDocumento4 páginasGametogénesis y ciclo sexual.docxValentina BOLDRINIAún no hay calificaciones
- ESPERMATOGENESISDocumento10 páginasESPERMATOGENESISAlfonso Quispe CopaAún no hay calificaciones
- Embriología: Desarrollo Ovulo Espermatozoide FecundaciónDocumento35 páginasEmbriología: Desarrollo Ovulo Espermatozoide FecundaciónLissetteAlexandraPerezAún no hay calificaciones
- GametogenesisDocumento6 páginasGametogenesisRodriguez KarlaAún no hay calificaciones
- La Sexualidad. Espermatogénesis y Ovogénesis.Documento3 páginasLa Sexualidad. Espermatogénesis y Ovogénesis.Aldair HernandezAún no hay calificaciones
- Células Germinales PrimordialesDocumento6 páginasCélulas Germinales PrimordialesLuis López Medina.Aún no hay calificaciones
- EspermatogénesisDocumento5 páginasEspermatogénesisXavier GuillenAún no hay calificaciones
- Embriología y Anatomia 1Documento27 páginasEmbriología y Anatomia 1Alejandro Fisac LanchaAún no hay calificaciones
- Lección No 1. Fisiología de La Reproduccion1Documento29 páginasLección No 1. Fisiología de La Reproduccion1vimarjuAún no hay calificaciones
- Cyd - Uabp 2 - Embrio - GametogenesisDocumento2 páginasCyd - Uabp 2 - Embrio - GametogenesisFlorencia BallardiniAún no hay calificaciones
- Gametognesis BiologiaDocumento15 páginasGametognesis BiologiaJosue RebolledoAún no hay calificaciones
- Ovogénesis y espermatogénesisDocumento10 páginasOvogénesis y espermatogénesisMaureenCatalinaSilvaLizanaAún no hay calificaciones
- GAMETOGÉNESIS - LangmanDocumento5 páginasGAMETOGÉNESIS - LangmanViviana PalaciosAún no hay calificaciones
- EMBRIOLOGIADocumento9 páginasEMBRIOLOGIAAlejandra MartinezAún no hay calificaciones
- T-6. MeiosisDocumento2 páginasT-6. MeiosisJuanJose45Aún no hay calificaciones
- GAMETOGENESIS ExposicionDocumento5 páginasGAMETOGENESIS ExposicionKEITLYN BENTHAMS SILVA100% (1)
- Tema 1 Desarrollo EmbrionarioDocumento19 páginasTema 1 Desarrollo EmbrionarioAnder GroundAún no hay calificaciones
- Gametogénesis y meiosis: La formación de gametosDocumento7 páginasGametogénesis y meiosis: La formación de gametosHeidy Juliana Ramírez GómezAún no hay calificaciones
- Cuadro Comparativo Entre Ovogenesis y EspermatogenesisDocumento5 páginasCuadro Comparativo Entre Ovogenesis y Espermatogenesisgavio_ta2669% (13)
- GametogénesisDocumento7 páginasGametogénesisMonics GonzalezAún no hay calificaciones
- Formación de gametos masculinos y femeninos (espermatozoides y óvulosDocumento9 páginasFormación de gametos masculinos y femeninos (espermatozoides y óvulosAixa CamposAún no hay calificaciones
- OvogénesisDocumento5 páginasOvogénesisKevin CisAún no hay calificaciones
- Embriología Resumen 1Documento20 páginasEmbriología Resumen 1Fidencio Jr.Aún no hay calificaciones
- Marco Teórico: Generalidades Del Aparato Reproductor MasculinoDocumento18 páginasMarco Teórico: Generalidades Del Aparato Reproductor MasculinoAndrea IriasAún no hay calificaciones
- Guia Gametogenesis, Fecundacion y 1ra SemanaDocumento5 páginasGuia Gametogenesis, Fecundacion y 1ra SemanaeldoctopuntocomAún no hay calificaciones
- Cambios Hormonales Asociados Al EmbarazoDocumento13 páginasCambios Hormonales Asociados Al EmbarazoLuisitaToroAún no hay calificaciones
- Esi 2Documento4 páginasEsi 2Nerina AndrikievichAún no hay calificaciones
- Guia Practicas Embriologia 2018Documento41 páginasGuia Practicas Embriologia 2018Wilson CCAún no hay calificaciones
- Ciclo Ovarico y Seg FolicularDocumento41 páginasCiclo Ovarico y Seg FolicularPierre GonzalesAún no hay calificaciones
- Patologias Del OvarioDocumento7 páginasPatologias Del OvarioKarina GoitiaAún no hay calificaciones
- Dibujo Blastocisto Día 7 - Mapa Conceptual - Dayana ChávezDocumento4 páginasDibujo Blastocisto Día 7 - Mapa Conceptual - Dayana ChávezDayana Anthonella Chavez MoralesAún no hay calificaciones
- Farmacologia EndocrinaDocumento42 páginasFarmacologia EndocrinaMaricela Ortiz Ochoa100% (1)
- OBSTETRICIA Y GINECOLOGÍA: Cambios fisiológicos, diagnóstico prenatalDocumento11 páginasOBSTETRICIA Y GINECOLOGÍA: Cambios fisiológicos, diagnóstico prenatalJuan Ramirez100% (1)
- Estudiar EmbriologiaDocumento14 páginasEstudiar EmbriologiaEVA YAMILETH FONSECA REYES100% (1)
- Lobo MexicanoDocumento12 páginasLobo MexicanoEduardo ReyesAún no hay calificaciones
- Embriología. Estructura y FunciónDocumento24 páginasEmbriología. Estructura y FunciónBenjamin Becerra FajardoAún no hay calificaciones
- Taller Del Ciclo Menstrual y Las Etapas de Formación de Un Bebé TALLER SIENCIAS NATURALEZ...Documento16 páginasTaller Del Ciclo Menstrual y Las Etapas de Formación de Un Bebé TALLER SIENCIAS NATURALEZ...deisyAún no hay calificaciones
- Apuntes ObstetriciaDocumento386 páginasApuntes ObstetriciaPedoraiemevalladolid100% (2)
- KUYENTUNDocumento16 páginasKUYENTUNhojas sonantes100% (2)
- Aula Quinto Grado A Octubre 1Documento12 páginasAula Quinto Grado A Octubre 1Any Lau LopezAún no hay calificaciones
- 3.anatomia Topografica 3.1 Posicion AnatomicaDocumento24 páginas3.anatomia Topografica 3.1 Posicion Anatomicacamila alvarezAún no hay calificaciones
- Potenciales de membrana y contracción muscularDocumento263 páginasPotenciales de membrana y contracción muscularErick Iñaguazo PindoAún no hay calificaciones
- Fisiología de La Pubertad MOD 2Documento5 páginasFisiología de La Pubertad MOD 2yoa_solizAún no hay calificaciones
- Fisiologia Reproductiva y Sistemas de Reproduccion de ConejosDocumento9 páginasFisiologia Reproductiva y Sistemas de Reproduccion de ConejosAmy MorejónAún no hay calificaciones
- Proceso de Formación Del HuevoDocumento2 páginasProceso de Formación Del HuevoVictor Franco SánchezAún no hay calificaciones
- Ciencia Y Tecnología: Comprobando Mis Logros V UnidadDocumento1 páginaCiencia Y Tecnología: Comprobando Mis Logros V UnidadRenato ZamudioAún no hay calificaciones
- Indicadores Naturales de La Ovulación y Bosas de ShawDocumento18 páginasIndicadores Naturales de La Ovulación y Bosas de ShawAnonymous ZnhRulAún no hay calificaciones
- Ensayo 6ta. J.E.G. PAES Anual-Biología 2023Documento50 páginasEnsayo 6ta. J.E.G. PAES Anual-Biología 2023oolaxd10Aún no hay calificaciones
- Parte de Renzoo-ReproducciónDocumento10 páginasParte de Renzoo-ReproducciónLizhyGalvanCalderonAún no hay calificaciones
- Sistema ReproductorDocumento21 páginasSistema ReproductorCieloAún no hay calificaciones
- Sincronizacion de Estros en Ganado BovinoDocumento44 páginasSincronizacion de Estros en Ganado BovinoEliseo CarpioAún no hay calificaciones
- 27 - 38Documento4 páginas27 - 38Taco Rex50% (2)
- Mecanismos de La MenstruacionDocumento15 páginasMecanismos de La MenstruacionMichael Cedeño LopezAún no hay calificaciones
- La Función de RelaciónDocumento10 páginasLa Función de RelaciónJennie McCoolAún no hay calificaciones
- Fisiología del ciclo menstrualDocumento4 páginasFisiología del ciclo menstrualfrankAún no hay calificaciones
- La conciencia en el cerebro: Descifrando el enigma de cómo el cerebro elabora nuestros pensamientosDe EverandLa conciencia en el cerebro: Descifrando el enigma de cómo el cerebro elabora nuestros pensamientosCalificación: 2.5 de 5 estrellas2.5/5 (3)
- En busca de la mente: El largo camino para comprender la vida mental (y lo que aún queda por descubrir)De EverandEn busca de la mente: El largo camino para comprender la vida mental (y lo que aún queda por descubrir)Calificación: 4.5 de 5 estrellas4.5/5 (3)
- Terapia cognitiva: Conceptos básicos y profundizaciónDe EverandTerapia cognitiva: Conceptos básicos y profundizaciónCalificación: 5 de 5 estrellas5/5 (1)
- Ansiedad infantil. Los trastornos explicados a los padresDe EverandAnsiedad infantil. Los trastornos explicados a los padresCalificación: 4.5 de 5 estrellas4.5/5 (25)
- El domador de cerebros: 40 propuestas para dominar tu mente y mejorar tu vidaDe EverandEl domador de cerebros: 40 propuestas para dominar tu mente y mejorar tu vidaCalificación: 4.5 de 5 estrellas4.5/5 (18)
- Terapia cognitiva del comportamiento: Descubre cómo despejar tu cerebro con la TCC. Aprender a superar el miedo y la ansiedad, la depresiónDe EverandTerapia cognitiva del comportamiento: Descubre cómo despejar tu cerebro con la TCC. Aprender a superar el miedo y la ansiedad, la depresiónCalificación: 5 de 5 estrellas5/5 (3)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- El autismo: Reflexiones y pautas para comprenderlo y abordarloDe EverandEl autismo: Reflexiones y pautas para comprenderlo y abordarloCalificación: 4 de 5 estrellas4/5 (7)
- Hipnoparto Y Parto Creativo: Cómo dar a luz con confianza y sin miedo Altravez del Método LeclaireDe EverandHipnoparto Y Parto Creativo: Cómo dar a luz con confianza y sin miedo Altravez del Método LeclaireCalificación: 4 de 5 estrellas4/5 (7)
- Cómo estimular el cerebro del niño: 100 ejercicios para potenciar la concentración, la memoria y otras funciones ejecutivasDe EverandCómo estimular el cerebro del niño: 100 ejercicios para potenciar la concentración, la memoria y otras funciones ejecutivasCalificación: 4.5 de 5 estrellas4.5/5 (12)
- Diccionario de neurociencias: Aplicadas a organizaciones y personasDe EverandDiccionario de neurociencias: Aplicadas a organizaciones y personasCalificación: 5 de 5 estrellas5/5 (4)
- Psicología y trastornos de los niños en la edad evolutiva: Qué son y cómo funcionanDe EverandPsicología y trastornos de los niños en la edad evolutiva: Qué son y cómo funcionanCalificación: 5 de 5 estrellas5/5 (1)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Neurociencia para vencer la depresión: La esprial ascendenteDe EverandNeurociencia para vencer la depresión: La esprial ascendenteCalificación: 4.5 de 5 estrellas4.5/5 (10)
- Anatomía del caballo: Guía práctica ilustradaDe EverandAnatomía del caballo: Guía práctica ilustradaCalificación: 4 de 5 estrellas4/5 (9)
- Autismo: guía para padres sobre el trastorno del espectro autista En españolDe EverandAutismo: guía para padres sobre el trastorno del espectro autista En españolCalificación: 4.5 de 5 estrellas4.5/5 (4)
- La comida y el inconsciente: Psicoanálisis y trastornos alimentariosDe EverandLa comida y el inconsciente: Psicoanálisis y trastornos alimentariosAún no hay calificaciones
- El cerebro matemático: Cómo nacen, viven y a veces mueren los números en nuestra menteDe EverandEl cerebro matemático: Cómo nacen, viven y a veces mueren los números en nuestra menteCalificación: 4 de 5 estrellas4/5 (5)
- Terapia Cognitivo Conductual: Psicología AplicadaDe EverandTerapia Cognitivo Conductual: Psicología AplicadaCalificación: 4 de 5 estrellas4/5 (5)
- Programa de la psicoeducación de la ansiedad y entrenamiento en técnicas de relajación: Manual del TerapeutaDe EverandPrograma de la psicoeducación de la ansiedad y entrenamiento en técnicas de relajación: Manual del TerapeutaCalificación: 4 de 5 estrellas4/5 (8)
- Casos clínicos: Evaluación, Diagnóstico e Intervención en Salud MentalDe EverandCasos clínicos: Evaluación, Diagnóstico e Intervención en Salud MentalCalificación: 5 de 5 estrellas5/5 (7)
- Trastorno por déficit de atención con hiperactividad: Una guía para padres y maestrosDe EverandTrastorno por déficit de atención con hiperactividad: Una guía para padres y maestrosCalificación: 5 de 5 estrellas5/5 (1)
- ¿Cómo aprendemos?: Los cuatro pilares con los que la educación puede potenciar los talentos de nuestro cerebroDe Everand¿Cómo aprendemos?: Los cuatro pilares con los que la educación puede potenciar los talentos de nuestro cerebroCalificación: 4 de 5 estrellas4/5 (14)
- Mindfulness: Cómo hacer menos y ser más conscientes puede ayudarnos a vivir mejor en este mundo estresadoDe EverandMindfulness: Cómo hacer menos y ser más conscientes puede ayudarnos a vivir mejor en este mundo estresadoCalificación: 4.5 de 5 estrellas4.5/5 (9)
- Neuromagia: Qué pueden enseñarnos los magos (y la ciencia) sobre el funcionamiento del cerebroDe EverandNeuromagia: Qué pueden enseñarnos los magos (y la ciencia) sobre el funcionamiento del cerebroCalificación: 4 de 5 estrellas4/5 (2)