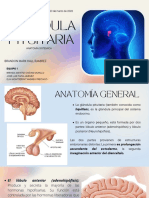
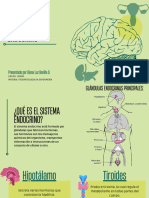
También podría gustarte
- El código de la obesidad: Descifrando los secretos de la pérdida de pesoDe EverandEl código de la obesidad: Descifrando los secretos de la pérdida de pesoCalificación: 4.5 de 5 estrellas4.5/5 (51)
- Transforma Tu CuerpoDocumento31 páginasTransforma Tu CuerpoDiana Cristina Mejia100% (1)
- Hipotiroidismo EDocumento39 páginasHipotiroidismo EMiquele Cannavaro50% (2)
- Gigantismo y Acromegalia - ScribdDocumento24 páginasGigantismo y Acromegalia - Scribdn4t4t4sAún no hay calificaciones
- Entrenamiento Concurrente de Fuerza y ResistenciaDocumento15 páginasEntrenamiento Concurrente de Fuerza y ResistenciaAlejandro IndaburuAún no hay calificaciones
- 04-Tejido AdiposoDocumento13 páginas04-Tejido Adiposojjoa4373395Aún no hay calificaciones
- 1 Hipotalamo Hipofisis-Completa 2017Documento49 páginas1 Hipotalamo Hipofisis-Completa 2017virginiaAún no hay calificaciones
- Poster - Hipertiroidismo - PDF 2Documento1 páginaPoster - Hipertiroidismo - PDF 2Juan Ricardo Sandoval SalcedoAún no hay calificaciones
- Proyecto de Tesis Obesidad Yaz y Mario ObesidadDocumento49 páginasProyecto de Tesis Obesidad Yaz y Mario ObesidadKatteryn QuimisAún no hay calificaciones
- Presentación Proyecto Científico Infantil Ilustrado Pastel Violeta y NaranjaDocumento14 páginasPresentación Proyecto Científico Infantil Ilustrado Pastel Violeta y NaranjaJeins PalominoAún no hay calificaciones
- CASO CLINICO Sistema Reproductor FemeninoDocumento5 páginasCASO CLINICO Sistema Reproductor FemeninoDavid Tiago Cueva50% (2)
- Glándula PituitariaDocumento14 páginasGlándula PituitariaYULIA ANDREA COHETO ALCOCERAún no hay calificaciones
- PC EndócrinoDocumento79 páginasPC EndócrinoAlan YañezAún no hay calificaciones
- Vol17 n2 para Saber de 01Documento7 páginasVol17 n2 para Saber de 01Andrey Jiménez CalderónAún no hay calificaciones
- Obesidad 2020Documento13 páginasObesidad 2020Maria de los Milagros JuárezAún no hay calificaciones
- 1guia 04 Biologia Grado OctavoDocumento3 páginas1guia 04 Biologia Grado OctavoJenny Vallinas PeñalozaAún no hay calificaciones
- Tema 17 EndoDocumento14 páginasTema 17 EndoUniversal UniversalAún no hay calificaciones
- Actualizacion en ObesidadDocumento8 páginasActualizacion en ObesidadadelaidaanguenvemengueAún no hay calificaciones
- Borrador de ObesidadDocumento3 páginasBorrador de Obesidadjuan topochoi mendezAún no hay calificaciones
- Sã Ndrome de Ovario Poliquã Stico 2022Documento24 páginasSã Ndrome de Ovario Poliquã Stico 2022Maria Ximena YáňezAún no hay calificaciones
- Cuestionario Solemne 2 Tecnología MédicaDocumento12 páginasCuestionario Solemne 2 Tecnología MédicaConstanza MendozaAún no hay calificaciones
- Obesidad, Anemia, Amenorrea y OsteomalaciaDocumento69 páginasObesidad, Anemia, Amenorrea y OsteomalaciaLaura Juliana Quintero WiderkehrAún no hay calificaciones
- Sistema EndocrinoDocumento8 páginasSistema EndocrinoMaríaAún no hay calificaciones
- Obesidad 2Documento40 páginasObesidad 2rodmigan89Aún no hay calificaciones
- Charla ObesidadDocumento34 páginasCharla ObesidadMaggie JacksonAún no hay calificaciones
- Infografia SopDocumento1 páginaInfografia SopXochitl hernandezAún no hay calificaciones
- Diario de Clase 8Documento6 páginasDiario de Clase 8Selene JumboAún no hay calificaciones
- CAPÍTULO 376 - Estudio Del Paciente Con Enfermedades EndocrinasDocumento8 páginasCAPÍTULO 376 - Estudio Del Paciente Con Enfermedades EndocrinasDaniela AlvarezAún no hay calificaciones
- Cedula de Hipotiroidismo CongenitoDocumento3 páginasCedula de Hipotiroidismo CongenitoRoxana Cruz AbarcaAún no hay calificaciones
- Síndrome de CushingDocumento6 páginasSíndrome de Cushingangel riosAún no hay calificaciones
- EndocrinologiaDocumento161 páginasEndocrinologiaSara DelgadoAún no hay calificaciones
- Enfermedades Crónicas No TransmisiblesDocumento28 páginasEnfermedades Crónicas No TransmisiblesLORENAAún no hay calificaciones
- 1.3 ObesidadDocumento6 páginas1.3 ObesidadFrida ParraAún no hay calificaciones
- Talla BajaDocumento22 páginasTalla BajaSamara JimenezAún no hay calificaciones
- Sistema EndocrinoDocumento36 páginasSistema EndocrinoGonzalo Álvarez MenéndezAún no hay calificaciones
- HipófisisDocumento14 páginasHipófisisMaria Laura vilchez ojedaAún no hay calificaciones
- Alteraciones en Hipotálamo e HipófisisDocumento15 páginasAlteraciones en Hipotálamo e HipófisisKarina GoitiaAún no hay calificaciones
- Patologías HipofisariasDocumento29 páginasPatologías Hipofisariassergio blancasAún no hay calificaciones
- Alteraciones EndocrinologicasDocumento50 páginasAlteraciones EndocrinologicasElizabeth Mondragon hoyosAún no hay calificaciones
- EndocrinoDocumento4 páginasEndocrinoSusan PeñaAún no hay calificaciones
- Alteraciones Del Ciclo SexualDocumento19 páginasAlteraciones Del Ciclo Sexualapi-26188232100% (1)
- Sindrome de MauriacDocumento3 páginasSindrome de MauriacAlejandro AponteAún no hay calificaciones
- ABP Glandula Suprarrenal GRUPO N°1Documento6 páginasABP Glandula Suprarrenal GRUPO N°1yo watchAún no hay calificaciones
- dIABETES TIPO I Y IIDocumento38 páginasdIABETES TIPO I Y IIClary SherwoodAún no hay calificaciones
- Sistema EndocrinoDocumento19 páginasSistema Endocrinolearnmore220Aún no hay calificaciones
- DM1 y SX de MauriacDocumento6 páginasDM1 y SX de MauriacViridiana Hernandez DiazAún no hay calificaciones
- Clase 12 y 13 - Hipofunción HipofisariaDocumento20 páginasClase 12 y 13 - Hipofunción HipofisariaUnidad de Apoyo a la Docencia e Investigación del HSJ-C100% (1)
- Caso Clinico Curso LasalleDocumento6 páginasCaso Clinico Curso LasalleClaudio R. MonteroAún no hay calificaciones
- Hipotiroidismo CongénitoDocumento4 páginasHipotiroidismo CongénitoBrandon UchihaAún no hay calificaciones
- Admin, V52n3a05Documento14 páginasAdmin, V52n3a05Analia LorensAún no hay calificaciones
- Enfermedades Del Sistema EndocrinoDocumento7 páginasEnfermedades Del Sistema EndocrinoLaura ChinchayAún no hay calificaciones
- 1.-Introducción Módulo de NutriciónDocumento46 páginas1.-Introducción Módulo de Nutriciónrodrigo roaAún no hay calificaciones
- Informe Legal DE FISIOLOGIADocumento15 páginasInforme Legal DE FISIOLOGIAJOS LUIS ALANOCA QUISPEAún no hay calificaciones
- Tema 2. Trastornos de La AdenohipofisisDocumento4 páginasTema 2. Trastornos de La AdenohipofisisMaria Bautista ContrerasAún no hay calificaciones
- Exposicion Introduccion HormonasDocumento8 páginasExposicion Introduccion HormonasValentina DiazAún no hay calificaciones
- 3 AnovulacionDocumento74 páginas3 AnovulacionEdgar EnriqueAún no hay calificaciones
- AcromegaliaDocumento40 páginasAcromegaliaDaniielita BeltranAún no hay calificaciones
- Obesidad - Grupo 3Documento11 páginasObesidad - Grupo 3Dennesis Katherine Peralta VeraAún no hay calificaciones
- Sistema EndocrinoDocumento6 páginasSistema Endocrinojuan francoAún no hay calificaciones
- Valoracionendocrinapropedeuticaii 091117154403 Phpapp01Documento22 páginasValoracionendocrinapropedeuticaii 091117154403 Phpapp01Gerald CrespinAún no hay calificaciones
- Clase XI - 2022 - Patología EndocrinaDocumento28 páginasClase XI - 2022 - Patología EndocrinaMaite RodriguezAún no hay calificaciones
- EndocrinoDocumento70 páginasEndocrinoGracestefi VieraAún no hay calificaciones
- OBESIDAD - Martha Beatriz González EspinoDocumento10 páginasOBESIDAD - Martha Beatriz González EspinoMartha HoranAún no hay calificaciones
- Diabetes UchileDocumento40 páginasDiabetes UchileEsteban MatusAún no hay calificaciones
- CAPÍTULO 408 - El Síndrome MetabólicoDocumento14 páginasCAPÍTULO 408 - El Síndrome Metabólicomaria joseAún no hay calificaciones
- LeptinaDocumento7 páginasLeptinaJose Angel Rojas MartinezAún no hay calificaciones
- Concepto de ObesidadDocumento26 páginasConcepto de ObesidadMario OliveraAún no hay calificaciones
- OBESIDADDocumento25 páginasOBESIDADEdgar Manuel Siguas Garcia0% (1)
- CAPÍTULO 401 - Biopatología de La ObesidadDocumento15 páginasCAPÍTULO 401 - Biopatología de La ObesidadGinecologia ObstetriciaAún no hay calificaciones
- Fisiología - Humana - (3a. - Ed.) - (PG - 1030 - 1208) - 107-118Documento12 páginasFisiología - Humana - (3a. - Ed.) - (PG - 1030 - 1208) - 107-118sthefanny gamezAún no hay calificaciones
- Nutricion Es Con-Ciencia PDFDocumento320 páginasNutricion Es Con-Ciencia PDFCAROL100% (1)
- Ayuno MáximoDocumento7 páginasAyuno MáximoGINA VANESA RAMIREZ ARAUJOAún no hay calificaciones
- La Energía de La VidaDocumento12 páginasLa Energía de La VidaOmeacatl MalitzinAún no hay calificaciones
- Tejido Adiposo PDF Clasificacion AnatomiaDocumento7 páginasTejido Adiposo PDF Clasificacion AnatomiaVikttoria Di DrtAún no hay calificaciones
- La LeptinaDocumento2 páginasLa LeptinaJorge Alexander Román MoralesAún no hay calificaciones
- Tejido Adiposo Como Organo Endocrino PDFDocumento9 páginasTejido Adiposo Como Organo Endocrino PDFManuel GonzalezAún no hay calificaciones
- Obesidad FINAL PDFDocumento41 páginasObesidad FINAL PDFSofia Machin0% (1)
- Amenorrea Hipotalámica FuncionalDocumento8 páginasAmenorrea Hipotalámica FuncionalmiguelalmenarezAún no hay calificaciones
- Ensayo Sobre La ObesidadDocumento26 páginasEnsayo Sobre La ObesidadyualitoAún no hay calificaciones
- Eje HHgonadal Adiposo, Leptina, Estrog, Anorexia, Bulimia, ObesidadDocumento6 páginasEje HHgonadal Adiposo, Leptina, Estrog, Anorexia, Bulimia, ObesidadMiguel De Bortoli100% (1)
- Relación Entre El Consumo Crónico de Heroína y Los Patrones AlimentariosDocumento54 páginasRelación Entre El Consumo Crónico de Heroína y Los Patrones AlimentariosMaux Fdz PardoAún no hay calificaciones
- CONSUMO VOLUNTARIO - Af.esDocumento8 páginasCONSUMO VOLUNTARIO - Af.esAna Sofía GarcesAún no hay calificaciones
- Pathogenesis and Treatment of Malnutrition in Patients On Maintenance Hemodialysis - UpToDateDocumento14 páginasPathogenesis and Treatment of Malnutrition in Patients On Maintenance Hemodialysis - UpToDateAlonso MedinaAún no hay calificaciones
- Odesidad y Sindrome MetabolicoDocumento29 páginasOdesidad y Sindrome MetabolicoLUZ ESMERALDA QUISPE MORENOAún no hay calificaciones
- Auriculoterapia Colombia PDFDocumento81 páginasAuriculoterapia Colombia PDFCata CastroAún no hay calificaciones
- CIRUGIA BARIATRICA Sabiston. Tratado de Cirugía 20 EdiciónDocumento24 páginasCIRUGIA BARIATRICA Sabiston. Tratado de Cirugía 20 EdiciónAnnery PaulinoAún no hay calificaciones
- Presentacion de Hormonas Antiestres en Los Salmones (Cortizol, Adrenalina)Documento22 páginasPresentacion de Hormonas Antiestres en Los Salmones (Cortizol, Adrenalina)Jean Carlos Motta OsmaAún no hay calificaciones
- Secrecion GastricaDocumento10 páginasSecrecion GastricaSebastian MendozaAún no hay calificaciones
- Consumo VoluntarioDocumento27 páginasConsumo VoluntarioMarielena GarciaAún no hay calificaciones
- Manual de Obesidad para Farmacéuticos Dietética y FarmacologíaDocumento182 páginasManual de Obesidad para Farmacéuticos Dietética y FarmacologíaEdgardo100% (3)