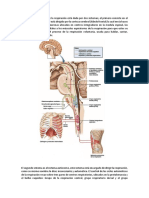
También podría gustarte
- 4 - Pancreas Endocrino PDFDocumento35 páginas4 - Pancreas Endocrino PDFteresa reyesAún no hay calificaciones
- DIABETESDocumento52 páginasDIABETESChristianAún no hay calificaciones
- 6 Pancreas EndocrinoDocumento53 páginas6 Pancreas EndocrinoFabiola ChadidAún no hay calificaciones
- Tema 8.5 Hormonas PancreáticasDocumento11 páginasTema 8.5 Hormonas Pancreáticaspruizi14Aún no hay calificaciones
- Pancreas, PDFDocumento110 páginasPancreas, PDFYesvicAún no hay calificaciones
- Clase 4 Pancreas EndocrinoDocumento28 páginasClase 4 Pancreas EndocrinoNelly Valencia CerveraAún no hay calificaciones
- Pancreatitis Aguda (Degraba 2019-II)Documento46 páginasPancreatitis Aguda (Degraba 2019-II)Kevin Quispe OchoaAún no hay calificaciones
- Teoria - GastrofisiologiaDocumento3 páginasTeoria - GastrofisiologiaAugusto Vargas FernandezAún no hay calificaciones
- Embarazo PatológicoDocumento45 páginasEmbarazo PatológicoBrandon Amanqui ApazaAún no hay calificaciones
- Itzel Tlecuitl Cerón-Pancreas EndocrinoDocumento3 páginasItzel Tlecuitl Cerón-Pancreas EndocrinoItzel Tlecuitl CerónAún no hay calificaciones
- E4 - Resumen Seminarios PDFDocumento15 páginasE4 - Resumen Seminarios PDFPau Funes GrovermanAún no hay calificaciones
- Sesión 6 - Pancreas Endocrino y Tejido AdiposoDocumento61 páginasSesión 6 - Pancreas Endocrino y Tejido AdiposoFa Fabi FabiiollaAún no hay calificaciones
- Manejo GlucosaDocumento23 páginasManejo GlucosaLex An DroAún no hay calificaciones
- DIABETESDocumento50 páginasDIABETESJuan Luis BalarezoAún no hay calificaciones
- Teoria 17Documento3 páginasTeoria 17IBRAHIM SEBASTIAN MENDOZA BAZANAún no hay calificaciones
- Clase EndocrinologiaDocumento221 páginasClase EndocrinologiajcarloscotuaAún no hay calificaciones
- Repaso Master de Embarazo Patológico Parte I 2022Documento45 páginasRepaso Master de Embarazo Patológico Parte I 2022juliana isolina calderon asencios100% (1)
- Diabetes Mellitus Tipo 1 y 2Documento85 páginasDiabetes Mellitus Tipo 1 y 2Melissa JustoAún no hay calificaciones
- Fisiología Capítulo 64Documento45 páginasFisiología Capítulo 64Anthony BorborAún no hay calificaciones
- Morfofisio PancreasDocumento2 páginasMorfofisio PancreasBenjamin HernandezAún no hay calificaciones
- PancreatitisDocumento63 páginasPancreatitisH.Melina Miranda RamirezAún no hay calificaciones
- Resumen Endocrino Dm1y2Documento3 páginasResumen Endocrino Dm1y2Abi HinojosaAún no hay calificaciones
- Cuadros DigestiónDocumento6 páginasCuadros DigestiónKcarck MviAún no hay calificaciones
- Páncreas EndocrinoDocumento78 páginasPáncreas EndocrinoKarla NoriegaAún no hay calificaciones
- Diabetes 2Documento9 páginasDiabetes 2Camila SanchezAún no hay calificaciones
- FSL Pancreas Endócrino ECopari-J 2019 PDFDocumento55 páginasFSL Pancreas Endócrino ECopari-J 2019 PDFjuan choqueAún no hay calificaciones
- Expo Karite Sin A Final 1Documento21 páginasExpo Karite Sin A Final 1Karina Curiel RamirezAún no hay calificaciones
- Anatomia y Fisiologia Del PáncreasDocumento26 páginasAnatomia y Fisiologia Del PáncreasSabelyAún no hay calificaciones
- Tumores Neuroendocrinos y GastroenteropancreáticosDocumento2 páginasTumores Neuroendocrinos y GastroenteropancreáticosPedroLinares94Aún no hay calificaciones
- Hormonas Del Pancreas y Del Tracto GastrointestinalDocumento21 páginasHormonas Del Pancreas y Del Tracto GastrointestinalChavez Flores Sergio IsmaelAún no hay calificaciones
- Practica 15Documento54 páginasPractica 15Rosario Sanchez SarmientoAún no hay calificaciones
- Absorción de Glucosa Intestinal Ultima VersiónDocumento22 páginasAbsorción de Glucosa Intestinal Ultima VersiónFreddy Rafael Altamirano ApanAún no hay calificaciones
- ENDOCRINOLOGIA 1 - EstudiosMyCDocumento144 páginasENDOCRINOLOGIA 1 - EstudiosMyCRenzoAún no hay calificaciones
- Integración y Regulación Metabólica: Dra. Gisela Pita Rodríguez Profesor AuxiliarDocumento42 páginasIntegración y Regulación Metabólica: Dra. Gisela Pita Rodríguez Profesor AuxiliarLilianEstherRuizGonzalezAún no hay calificaciones
- DownloadDocumento14 páginasDownloadGino Eduardo Taboada LozanoAún no hay calificaciones
- Guia para El Manejo de La Insulina en La Diabetes Tipo 2Documento17 páginasGuia para El Manejo de La Insulina en La Diabetes Tipo 2David Alvarado MartínezAún no hay calificaciones
- AMBOSS FisiologíaDocumento27 páginasAMBOSS FisiologíaRonald Alexander Campoverde RiosAún no hay calificaciones
- Diabetes Mellitus Ver Alumno 2020Documento73 páginasDiabetes Mellitus Ver Alumno 2020Paola Rodríguez GuzmánAún no hay calificaciones
- Hormonas Gastrointestinales 2.0Documento26 páginasHormonas Gastrointestinales 2.0Sianya Gutierrez100% (1)
- Trastornos Relacionados Con La Secreción Gástrica ÁcidaDocumento26 páginasTrastornos Relacionados Con La Secreción Gástrica ÁcidaValeria Arauz SubiagaAún no hay calificaciones
- Glándulas Anexas 2024 - PDFDocumento11 páginasGlándulas Anexas 2024 - PDFMatias DelgadoAún no hay calificaciones
- PáncreasDocumento10 páginasPáncreasDiana Noeli Ramirez IbañezAún no hay calificaciones
- Fisiología Del Aparato DigestivoDocumento46 páginasFisiología Del Aparato DigestivoKeniia De La VegaAún no hay calificaciones
- Temas 3er Parcial PEDIATRIA IDocumento49 páginasTemas 3er Parcial PEDIATRIA IDarvin GarcíaAún no hay calificaciones
- Fisiopatología de La Diabetes MellitusDocumento51 páginasFisiopatología de La Diabetes MellitusGabriela GarcíaAún no hay calificaciones
- Fisio Gastro ResueltoDocumento44 páginasFisio Gastro ResueltoCielo Perez MendozaAún no hay calificaciones
- Histologia Células Del Estómago y Su FunciónDocumento3 páginasHistologia Células Del Estómago y Su FunciónKrystel Volgger Garcia100% (2)
- Pruebas de La Función RenalDocumento37 páginasPruebas de La Función RenalFranklin Tacuchi JaraAún no hay calificaciones
- Diarrea en PediatríaDocumento30 páginasDiarrea en PediatríaSUSANA RAQUEL GUARDADO GOMEZAún no hay calificaciones
- Fisiología Endócrina Del Páncreas. Dra. Gabriela LoyaDocumento45 páginasFisiología Endócrina Del Páncreas. Dra. Gabriela LoyaJairJean Zevallos GuereAún no hay calificaciones
- Biblio 4 TraducidoDocumento6 páginasBiblio 4 TraducidoDenise Da Silva GoncalvesAún no hay calificaciones
- Diabetes Mellitus Wiener 2020-1Documento107 páginasDiabetes Mellitus Wiener 2020-1Medalyt Huashuayo CusiAún no hay calificaciones
- Anato Fisio PancreasDocumento63 páginasAnato Fisio Pancreasfranciscomejia14835Aún no hay calificaciones
- Pancreatitis AgudaDocumento10 páginasPancreatitis AgudaValentina AndradeAún no hay calificaciones
- Control de GlucemiaDocumento4 páginasControl de GlucemiaSarahCardosoAún no hay calificaciones
- Enzimología ClínicaDocumento4 páginasEnzimología Clínicalucia marquez lopezAún no hay calificaciones
- TnegepDocumento1 páginaTnegepJose Cordero (PianoGuyJD)Aún no hay calificaciones
- CUADRO HipoglucemiantesDocumento3 páginasCUADRO HipoglucemiantesVinicius AlvesAún no hay calificaciones
- Homeostasis Renal de GlucosaDocumento37 páginasHomeostasis Renal de GlucosaJosé Guilherme Batista RodriguesAún no hay calificaciones
- Shigella Sp.Documento10 páginasShigella Sp.Guadalupe Mendoza RadasAún no hay calificaciones
- Algoritmo NacDocumento4 páginasAlgoritmo NacGuadalupe Mendoza RadasAún no hay calificaciones
- INFORME Cándida AlbicansDocumento13 páginasINFORME Cándida AlbicansGuadalupe Mendoza RadasAún no hay calificaciones
- Proteína C ReactivaDocumento7 páginasProteína C ReactivaGuadalupe Mendoza RadasAún no hay calificaciones
- Caso Clínico GalactorreaDocumento2 páginasCaso Clínico GalactorreaGuadalupe Mendoza RadasAún no hay calificaciones
- Regulación de La RespiraciónDocumento3 páginasRegulación de La RespiraciónGuadalupe Mendoza RadasAún no hay calificaciones
- Casos ClinicosDocumento1 páginaCasos ClinicosGuadalupe Mendoza RadasAún no hay calificaciones
- Obtención de Animales TransgénicosDocumento11 páginasObtención de Animales TransgénicosVictorino KjAún no hay calificaciones
- Microbiologia Stanchi PDFDocumento588 páginasMicrobiologia Stanchi PDFelizabeth100% (1)
- Unit 1 - TAREADocumento5 páginasUnit 1 - TAREALuis CervantesAún no hay calificaciones
- Infografía de La PlantaDocumento22 páginasInfografía de La PlantaElkin PenagosAún no hay calificaciones
- Lenguaje Médico: Epónimos, Abreviaturas y Terminología Médica de Uso Frecuente.Documento14 páginasLenguaje Médico: Epónimos, Abreviaturas y Terminología Médica de Uso Frecuente.RoyserAún no hay calificaciones
- Resumen Sociologia DerechoDocumento25 páginasResumen Sociologia DerechoAgusSabatiniAún no hay calificaciones
- Triquinosis, TeniosisDocumento8 páginasTriquinosis, TeniosisAdriana Loayza PeraltaAún no hay calificaciones
- Etiologc3ada de La Enfermedad Periodontal PDFDocumento124 páginasEtiologc3ada de La Enfermedad Periodontal PDFVictor CrAún no hay calificaciones
- Neurociencia y Conducta I PDFDocumento11 páginasNeurociencia y Conducta I PDFOscar AlvarezAún no hay calificaciones
- Tratamiento Con PRP PDFDocumento9 páginasTratamiento Con PRP PDFMatias ArdilaAún no hay calificaciones
- Cuadro Comparativo Mitosis y MeiosisDocumento4 páginasCuadro Comparativo Mitosis y MeiosisIván Esquivel0% (1)
- Anemias RegenerativasDocumento154 páginasAnemias RegenerativaswillmedAún no hay calificaciones
- Circulacion FetalDocumento14 páginasCirculacion FetalGonzalez NanyAún no hay calificaciones
- Influencia de La Luz en La Produccion deDocumento8 páginasInfluencia de La Luz en La Produccion deBalamAún no hay calificaciones
- Guia Ciencias Funcion de Relacion en Las Plantas Profe LudiviaDocumento2 páginasGuia Ciencias Funcion de Relacion en Las Plantas Profe LudiviaGerson Leandro Rizo100% (1)
- Informe Biologia (Tejidos)Documento24 páginasInforme Biologia (Tejidos)Anderson Vargas0% (2)
- Control Microbiologico de AmbientesDocumento11 páginasControl Microbiologico de AmbientesLV LMAún no hay calificaciones
- Teoria Cromosomica de La HerenciaDocumento5 páginasTeoria Cromosomica de La HerenciaRoseAún no hay calificaciones
- Manual de Neurociencia Aplicada A La Educacion 2013 UDDDocumento105 páginasManual de Neurociencia Aplicada A La Educacion 2013 UDDEsteban Oni Dinamarca Huenulef100% (1)
- 1 Fisiologia de La Sangre PDFDocumento153 páginas1 Fisiologia de La Sangre PDFSalvatore AmezquitaAún no hay calificaciones
- Sopas de LetrasDocumento14 páginasSopas de Letrasclapacha.pAún no hay calificaciones
- Bio CompostDocumento7 páginasBio CompostErick Martinez LeónAún no hay calificaciones
- Historieta de NeuronaDocumento1 páginaHistorieta de NeuronaMiguel Angel Hernandez ArteagaAún no hay calificaciones
- 7 Principales Corrientes Teóricas de La AntropologíaDocumento61 páginas7 Principales Corrientes Teóricas de La AntropologíaJose WCAún no hay calificaciones
- Virus Mapa ConceptualDocumento1 páginaVirus Mapa ConceptualRosy Cachay LojaAún no hay calificaciones
- Banco de PreguntasDocumento215 páginasBanco de PreguntasjosemerelloAún no hay calificaciones
- Solucionario CT Primaria MAESTROS PERUANOS 26 NOVDocumento56 páginasSolucionario CT Primaria MAESTROS PERUANOS 26 NOVluis tu100% (1)
- Cdi U1 Ea ApggDocumento7 páginasCdi U1 Ea ApggApolo García García50% (4)
- Conferencia 11 Leyes de MendelDocumento8 páginasConferencia 11 Leyes de MendelLISBETH DENNISSE CALE�O MONTEROAún no hay calificaciones
- Células FusiformesDocumento4 páginasCélulas FusiformespeluchinsiAún no hay calificaciones