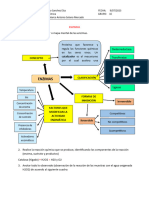
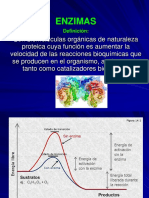
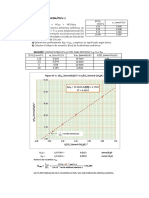
También podría gustarte
- Enzimas: Características y ClasificaciónDocumento27 páginasEnzimas: Características y ClasificaciónvictorAún no hay calificaciones
- Enzimas y vitaminasDocumento28 páginasEnzimas y vitaminasAdelfa BonalumiAún no hay calificaciones
- Usmp EnzimasDocumento56 páginasUsmp Enzimasapi-3708303100% (5)
- EnzimasDocumento29 páginasEnzimasmanu92egAún no hay calificaciones
- Taller de EnzimasDocumento5 páginasTaller de EnzimasivanniaAún no hay calificaciones
- 08 EnzimasDocumento44 páginas08 EnzimasCarlos M. Guerra AAún no hay calificaciones
- Metabolismo CelularDocumento9 páginasMetabolismo CelularamandaAún no hay calificaciones
- Enzimas NuevoDocumento41 páginasEnzimas NuevoCardemio VillalobosAún no hay calificaciones
- Catalizadores y EnzimasDocumento24 páginasCatalizadores y EnzimasElias BoychukAún no hay calificaciones
- Enzimas: catalizadores biológicosDocumento145 páginasEnzimas: catalizadores biológicosriiAún no hay calificaciones
- Enzimas en Ing. de Bioprocesos PDFDocumento145 páginasEnzimas en Ing. de Bioprocesos PDFandesonAún no hay calificaciones
- TEMA 2 BIOQUIMICADocumento12 páginasTEMA 2 BIOQUIMICAMarta Moreno FrescoAún no hay calificaciones
- Bioenergética y transporte celularDocumento12 páginasBioenergética y transporte celularNicole Jensen100% (1)
- Bitacora 4 Enzimas Biologia CelularDocumento8 páginasBitacora 4 Enzimas Biologia CelularCortés Ramos Yessica SurikeyAún no hay calificaciones
- Informe EnzimasDocumento4 páginasInforme EnzimasElzy PeredoAún no hay calificaciones
- Unidad 3 EnzimasDocumento42 páginasUnidad 3 EnzimasGar Ara100% (1)
- Catalizadores Organicos 2022Documento39 páginasCatalizadores Organicos 2022Catherin HidalgoAún no hay calificaciones
- Taller de Bioquimica PDFDocumento12 páginasTaller de Bioquimica PDFjuan jose montalvoAún no hay calificaciones
- BIOQUIMICA CLASE REBECADocumento8 páginasBIOQUIMICA CLASE REBECArgarcesrendonAún no hay calificaciones
- EnzimasDocumento23 páginasEnzimasMaruby MontonerAún no hay calificaciones
- ENZIMASDocumento38 páginasENZIMASSr FaVAún no hay calificaciones
- Taller de EnzimasDocumento6 páginasTaller de Enzimasalexander acosta solanoAún no hay calificaciones
- Las Enzimas PDFDocumento9 páginasLas Enzimas PDFTriny VilladiegoAún no hay calificaciones
- 6 EnzimasDocumento52 páginas6 Enzimasgianluca malerbaAún no hay calificaciones
- EnzimasDocumento15 páginasEnzimasDavinia García MolinaAún no hay calificaciones
- Enzimas - Reaccion EnzimaticaDocumento65 páginasEnzimas - Reaccion EnzimaticaLizet Mayli Mejia Penadillo100% (2)
- ENZIMASDocumento39 páginasENZIMASDanilo Román AspilcuetaAún no hay calificaciones
- Catálisis EnzimáticaDocumento54 páginasCatálisis EnzimáticaNadiaAún no hay calificaciones
- Bioquimica Nutricional Enzimas-Tema 2Documento56 páginasBioquimica Nutricional Enzimas-Tema 2Ana MedinaAún no hay calificaciones
- Folleto Exposicion Catalisis EnzimaticaDocumento3 páginasFolleto Exposicion Catalisis EnzimaticaJohan CortesAún no hay calificaciones
- Apuntes de EnzimasDocumento4 páginasApuntes de EnzimasYadira Ramirez ReyesAún no hay calificaciones
- ENZIMASDocumento22 páginasENZIMASlauraAún no hay calificaciones
- Tema 3 ENZIMASDocumento145 páginasTema 3 ENZIMASPaulina Campoy RamirezAún no hay calificaciones
- Biocatalizadores y Referencias BibliograficasDocumento6 páginasBiocatalizadores y Referencias BibliograficasMaria Elizabeth M. Alberca0% (1)
- Biología Torado (08.54)Documento28 páginasBiología Torado (08.54)Impresiones CBCAún no hay calificaciones
- Serie Cinética Pérez Hernández Víctor ManuelDocumento2 páginasSerie Cinética Pérez Hernández Víctor ManuelVíctor Manuel Pérez HernándezAún no hay calificaciones
- En ZimasoDocumento34 páginasEn ZimasoKevin Rosas vasquezAún no hay calificaciones
- Bioquimica - Era IDocumento66 páginasBioquimica - Era IRoAún no hay calificaciones
- EnzimasDocumento34 páginasEnzimasBelen Del Rio100% (2)
- El Metabolismo y EnzimasDocumento42 páginasEl Metabolismo y EnzimasGuillermo ChavesAún no hay calificaciones
- En ZimasDocumento16 páginasEn ZimasErickAlmoraxdAncasiAún no hay calificaciones
- EnzimasDocumento12 páginasEnzimasmakoki92Aún no hay calificaciones
- Tema 3Documento9 páginasTema 3Erika Luna JiménezAún no hay calificaciones
- Ejercicio3 - Tarea 2 - Grupo Colaborativo 201103 - 65Documento26 páginasEjercicio3 - Tarea 2 - Grupo Colaborativo 201103 - 65Luisa Gil EAún no hay calificaciones
- Enzimas2023 CHDocumento25 páginasEnzimas2023 CHconsueloAún no hay calificaciones
- Apuntes Clase 2 BiocatalisisDocumento3 páginasApuntes Clase 2 Biocatalisisyasmin zavaleta velezAún no hay calificaciones
- Modelos de Acción EnzimáticaDocumento5 páginasModelos de Acción Enzimáticaluana ayalaAún no hay calificaciones
- EnzimasDocumento61 páginasEnzimasGeraldine Huarcaya Olivera100% (1)
- 3-Catálisis Enzimática ResumenDocumento9 páginas3-Catálisis Enzimática Resumenninas50544Aún no hay calificaciones
- Enzimas: definición, estructura, clasificación y funciónDocumento53 páginasEnzimas: definición, estructura, clasificación y funciónNelson BeltranAún no hay calificaciones
- Catalizadores EnzimáticosDocumento15 páginasCatalizadores EnzimáticosAlembert ZapataAún no hay calificaciones
- Taller de Las EnzimasDocumento10 páginasTaller de Las EnzimasJuan CamiloAún no hay calificaciones
- Enzimas y vitaminas: estructura, acción y regulaciónDocumento41 páginasEnzimas y vitaminas: estructura, acción y regulaciónstiben novaAún no hay calificaciones
- 5 - EnzimasDocumento7 páginas5 - Enzimasnahiarabeluu2Aún no hay calificaciones
- Enzimas Jd.Documento8 páginasEnzimas Jd.MaryLaderaMagoAún no hay calificaciones
- Enzimas BioquimicaDocumento45 páginasEnzimas BioquimicaJcaminoAún no hay calificaciones
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- Química de macrocomponentes de alimentosDe EverandQuímica de macrocomponentes de alimentosAún no hay calificaciones
- Problema 13.5-WalpoleDocumento4 páginasProblema 13.5-WalpoleEmmanuel RomanAún no hay calificaciones
- Enzimas, clasificación, estructura y factores que afectan su actividadDocumento1 páginaEnzimas, clasificación, estructura y factores que afectan su actividadMonik ReyesAún no hay calificaciones
- Choice Sobre Hidratos de CarbonoDocumento3 páginasChoice Sobre Hidratos de CarbonoJack MondragonAún no hay calificaciones
- Estructura: Transporte Activo: El ATP Desempeña Un Papel Crítico en ElDocumento3 páginasEstructura: Transporte Activo: El ATP Desempeña Un Papel Crítico en ElMitzi Estrada Alamo100% (1)
- TRANSCRIPCIONDocumento47 páginasTRANSCRIPCIONAlexa GarroniAún no hay calificaciones
- 2.1. CofactoresDocumento28 páginas2.1. CofactoresGigi AguiarAún no hay calificaciones
- BIOMOLÉCULASDocumento38 páginasBIOMOLÉCULASGeronimo PoetAún no hay calificaciones
- Inmunoglobulina ADocumento13 páginasInmunoglobulina AIrving MayoAún no hay calificaciones
- Tema 5 - AaDocumento8 páginasTema 5 - AaMarina Los LópezAún no hay calificaciones
- Temario Especifico (Alfredo) - Tema 12 - T-12 - Alimentacion-Nutricion PDFDocumento233 páginasTemario Especifico (Alfredo) - Tema 12 - T-12 - Alimentacion-Nutricion PDFEric2202Aún no hay calificaciones
- EL GENOMA DE UNA CÉLULA y SISTEMAS ENERGETICOS SUBCELULARESDocumento5 páginasEL GENOMA DE UNA CÉLULA y SISTEMAS ENERGETICOS SUBCELULARESRodrigoMendozaAún no hay calificaciones
- Las MitocondriasDocumento2 páginasLas Mitocondrias0ZKRCITHOAún no hay calificaciones
- REACCIONES ENZIMÁTICAS DE ÓXIDOREDUCCIÓNDocumento7 páginasREACCIONES ENZIMÁTICAS DE ÓXIDOREDUCCIÓNVale CarBoAún no hay calificaciones
- RabanoDocumento20 páginasRabanomichelAún no hay calificaciones
- ADN replicación y expresión génica tallerDocumento3 páginasADN replicación y expresión génica tallerErikaBedoya0% (1)
- Cascada de CoagulaciónDocumento9 páginasCascada de CoagulaciónLoezaAbiAún no hay calificaciones
- Enzimas DesarrolladoDocumento21 páginasEnzimas DesarrolladoweikeAún no hay calificaciones
- Arancel 2019 Servicios LaboratorioDocumento15 páginasArancel 2019 Servicios Laboratorionessa castilloAún no hay calificaciones
- Insulina CLLDocumento19 páginasInsulina CLLrafael mendozaAún no hay calificaciones
- Macromoléculas EnsayoDocumento5 páginasMacromoléculas EnsayoAndres MedinaAún no hay calificaciones
- ¿Hay Aminoácidos Azufrados en Las Proteínas de La Clara de HuevoDocumento1 página¿Hay Aminoácidos Azufrados en Las Proteínas de La Clara de HuevoSheila CruzAún no hay calificaciones
- Catabolismo de AminoacidosDocumento20 páginasCatabolismo de AminoacidosXimena Véliz DaudAún no hay calificaciones
- Metabolismo Del Paciente QuirúrgicoDocumento27 páginasMetabolismo Del Paciente QuirúrgicoMaxwell Enmanuel Hernández CentenoAún no hay calificaciones
- REPORTE Codigo GeneticoDocumento7 páginasREPORTE Codigo GeneticoLiliBolanos100% (1)
- Trabajo de Investigación de Aminoácidos y ProteínasDocumento40 páginasTrabajo de Investigación de Aminoácidos y ProteínasKRISHNA MICHELLE RODRIGUEZ FERNANDEZAún no hay calificaciones
- Guía Nº4 MacromoléculasDocumento17 páginasGuía Nº4 MacromoléculascamiloAún no hay calificaciones
- Cinetica Enzimatica 1Documento2 páginasCinetica Enzimatica 1Massiel Marley LOPEZ RODRIGUEZAún no hay calificaciones
- Producción de Especies Reactivas de Oxígeno por Células del Sistema InmuneDocumento20 páginasProducción de Especies Reactivas de Oxígeno por Células del Sistema InmuneGibran Padrón GuzmánAún no hay calificaciones
- Glutation PrometeoDocumento1 páginaGlutation PrometeoJosué EsparzaAún no hay calificaciones