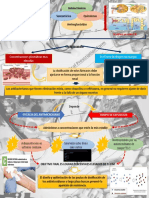
También podría gustarte
- Sepsis: Fisiopatología, diagnóstico y tratamientoDe EverandSepsis: Fisiopatología, diagnóstico y tratamientoAún no hay calificaciones
- Respuesta Inflamatoria Aguda, Consideraciones Bioquímicas y CelularesDocumento16 páginasRespuesta Inflamatoria Aguda, Consideraciones Bioquímicas y CelularesJorge Sanchez UlloaAún no hay calificaciones
- .Respuesta Inflamatoria Aguda. Consideraciones Bioquímicas y CelularesDocumento16 páginas.Respuesta Inflamatoria Aguda. Consideraciones Bioquímicas y CelularesFelipe Montiel MirandaAún no hay calificaciones
- Respuesta Inflamatoria AgudaDocumento3 páginasRespuesta Inflamatoria AgudaGonzález Almazán Vetzaira EneydaAún no hay calificaciones
- Bases 1Documento9 páginasBases 1Barbara AzevedoAún no hay calificaciones
- Jimenez Chavez Luis Fernando-3b-Iesch Medicina-Procesos Inmunologicos - Actividad 1Documento23 páginasJimenez Chavez Luis Fernando-3b-Iesch Medicina-Procesos Inmunologicos - Actividad 1Fernan lucasAún no hay calificaciones
- Agentes InflamatoriosDocumento15 páginasAgentes InflamatoriosagnikaiAún no hay calificaciones
- Agabriela - Unidad 1 - Actividad 2 - Concepto de Inflamación, Clasificación y Características GeneralesDocumento8 páginasAgabriela - Unidad 1 - Actividad 2 - Concepto de Inflamación, Clasificación y Características Generalesdaniel chaparroAún no hay calificaciones
- Resumen PatologiaDocumento16 páginasResumen PatologiaEmanuel D SangregorioAún no hay calificaciones
- INFLAMACIÓNDocumento12 páginasINFLAMACIÓNfanny2609Aún no hay calificaciones
- Agentes InflamatoriosDocumento19 páginasAgentes InflamatoriosRichard Leonidas P. LozaAún no hay calificaciones
- Trauma y SepsisDocumento12 páginasTrauma y SepsisNutriólogo Adar LazaroAún no hay calificaciones
- InflamaciónDocumento16 páginasInflamaciónSindy Cascante100% (1)
- INFLAMACIONDocumento7 páginasINFLAMACIONCintia Ortiz llanosAún no hay calificaciones
- Apunte INFLAMACION REPARACIONDocumento41 páginasApunte INFLAMACION REPARACIONAntonia LobosAún no hay calificaciones
- Inflamación Aguda y CrónicaDocumento44 páginasInflamación Aguda y CrónicaVioleta ChitayAún no hay calificaciones
- Capitulo 2Documento40 páginasCapitulo 2Jose Antonio Marrasquin LujanoAún no hay calificaciones
- InflamaciónDocumento11 páginasInflamaciónAbner Hernandez LopezAún no hay calificaciones
- Reactantes de Fase Aguda - Rol Proceso InflamatorioDocumento10 páginasReactantes de Fase Aguda - Rol Proceso InflamatorioCatalina GaticaAún no hay calificaciones
- Inflamacion y Reparacion PDFDocumento23 páginasInflamacion y Reparacion PDFSofiMuñozAún no hay calificaciones
- Inflamacion y Reparacion TisularDocumento18 páginasInflamacion y Reparacion TisularGenezareth Araya JulioAún no hay calificaciones
- Inflamacion Aguda y Cambios VascularesDocumento7 páginasInflamacion Aguda y Cambios VascularesIlliams ZetaAún no hay calificaciones
- Monografia de InflamacionDocumento16 páginasMonografia de InflamacionRichard Salazar RequenaAún no hay calificaciones
- Procesos InflamatoriosDocumento9 páginasProcesos InflamatoriosRenAud HeleneAún no hay calificaciones
- Unidad V Inflamacion Aguda y CronicaDocumento83 páginasUnidad V Inflamacion Aguda y CronicaBryan TalaveraAún no hay calificaciones
- 3 Respuesta Metabolica Al TraumaDocumento11 páginas3 Respuesta Metabolica Al TraumaDavid MayoAún no hay calificaciones
- Inflamación Equipo5Documento20 páginasInflamación Equipo5Layna RiAún no hay calificaciones
- 3 - Inflamacion Aguda REVISADADocumento13 páginas3 - Inflamacion Aguda REVISADASebastián Esteban González CarrascoAún no hay calificaciones
- Lectura Proceso - InflamatorioDocumento7 páginasLectura Proceso - Inflamatorioeudicsalazar2Aún no hay calificaciones
- Exposición - Equipo 9 LLDocumento13 páginasExposición - Equipo 9 LLÁngel LagunasAún no hay calificaciones
- INFLAMACIÓN - CirugiaDocumento7 páginasINFLAMACIÓN - CirugiaSadi danielaAún no hay calificaciones
- InflamacionDocumento5 páginasInflamacionenrique natarenAún no hay calificaciones
- InflamaciónDocumento8 páginasInflamaciónJhon HortealesAún no hay calificaciones
- Guia de Inflamacion y Reparacion Tisular Ua 2017Documento17 páginasGuia de Inflamacion y Reparacion Tisular Ua 2017Juan Pablo LagosAún no hay calificaciones
- Proceso InflamatorioDocumento3 páginasProceso InflamatoriosusanAún no hay calificaciones
- Inflamación y Reparación TerminadoDocumento29 páginasInflamación y Reparación TerminadoAxel Nuñez100% (2)
- Inflamacion Aguda y CrónicaDocumento13 páginasInflamacion Aguda y CrónicaEmilio BrolloAún no hay calificaciones
- Inflamación y Reparación TisularDocumento24 páginasInflamación y Reparación TisularCarolina Martinez Montano100% (1)
- Síndrome de ReperfusiónDocumento9 páginasSíndrome de ReperfusiónFernando Gonzalez MtzAún no hay calificaciones
- Síndrome de Respuesta Inflamatoria SistémicaDocumento6 páginasSíndrome de Respuesta Inflamatoria Sistémicalucero166Aún no hay calificaciones
- 001 Sris y SdmoDocumento6 páginas001 Sris y Sdmovitorinno5705100% (2)
- InflamacionDocumento37 páginasInflamacionRuth ZuritaAún no hay calificaciones
- SepsisDocumento12 páginasSepsisOscar FatushAún no hay calificaciones
- Plan de Clase MFPH I. AO 6Documento11 páginasPlan de Clase MFPH I. AO 6AngelAún no hay calificaciones
- InflamacionDocumento16 páginasInflamacionPriscilaqj 2001Aún no hay calificaciones
- INFLAMACIÓN para El ExamenDocumento10 páginasINFLAMACIÓN para El ExamenStephanieAún no hay calificaciones
- Inflamación ProDocumento19 páginasInflamación ProDaillynAún no hay calificaciones
- Patología de La InflamaciónDocumento5 páginasPatología de La InflamaciónLizeth GarciaAún no hay calificaciones
- Monografia InflamacionDocumento27 páginasMonografia InflamacionJudithGonzálesInga100% (2)
- Inflamación y SIRS - Unidad IIDocumento27 páginasInflamación y SIRS - Unidad IIMartin Inmediato GhettiAún no hay calificaciones
- Fisiopatologia de La InflamacionDocumento8 páginasFisiopatologia de La InflamacionCarla100% (1)
- InflamaciónDocumento8 páginasInflamaciónadriana aguileraAún no hay calificaciones
- Inflamacion RobbinsDocumento14 páginasInflamacion RobbinsAnnie ElizabethAún no hay calificaciones
- TP N2 Inflamacion AgudaDocumento12 páginasTP N2 Inflamacion AgudaLuis Ariel Rocha RodríguezAún no hay calificaciones
- TEORICADocumento9 páginasTEORICAQuenta Luque DavidAún no hay calificaciones
- Barrera - Rafael - Concepto de Inflamación, Clasificación y Características GeneralesDocumento10 páginasBarrera - Rafael - Concepto de Inflamación, Clasificación y Características GeneralesJOACHINAún no hay calificaciones
- Natural Resolution of Inflammation - En.es PDFDocumento16 páginasNatural Resolution of Inflammation - En.es PDFMafe TierradentroAún no hay calificaciones
- Inflamación Aguda, Cronica y Reparación TisularDocumento9 páginasInflamación Aguda, Cronica y Reparación TisularMariaAún no hay calificaciones
- General de La Inflamación y La Reparación Tisular e Inflamacion AgudaDocumento35 páginasGeneral de La Inflamación y La Reparación Tisular e Inflamacion AgudaJennifer SamanthaAún no hay calificaciones
- Semn 11Documento2 páginasSemn 11Jackelin Jhessabel Quino MenesesAún no hay calificaciones
- Elaborado y RevisadoDocumento1 páginaElaborado y RevisadoJackelin Jhessabel Quino MenesesAún no hay calificaciones
- Grupo 1 - Sistema RespiratorioDocumento12 páginasGrupo 1 - Sistema RespiratorioJackelin Jhessabel Quino MenesesAún no hay calificaciones
- Las Condiciones DesencadenantesDocumento2 páginasLas Condiciones DesencadenantesJackelin Jhessabel Quino MenesesAún no hay calificaciones
- Trabajo Grupal 1Documento14 páginasTrabajo Grupal 1Jackelin Jhessabel Quino MenesesAún no hay calificaciones
- Paginas VisistadasDocumento1 páginaPaginas VisistadasJackelin Jhessabel Quino MenesesAún no hay calificaciones
- Electroquímica: Tabla de Potenciales de CeldaDocumento2 páginasElectroquímica: Tabla de Potenciales de CeldaJackelin Jhessabel Quino MenesesAún no hay calificaciones
- Horario de EstudiosDocumento2 páginasHorario de EstudiosJackelin Jhessabel Quino MenesesAún no hay calificaciones
- Módulo Fisiopatología 2022-IIDocumento117 páginasMódulo Fisiopatología 2022-IIJackelin Jhessabel Quino MenesesAún no hay calificaciones
- Caso ClinicoDocumento5 páginasCaso ClinicoJackelin Jhessabel Quino MenesesAún no hay calificaciones
- HGVLJHGVBDocumento21 páginasHGVLJHGVBJackelin Jhessabel Quino MenesesAún no hay calificaciones
- Examen Final de Ingles Ultima SemanaDocumento1 páginaExamen Final de Ingles Ultima SemanaJackelin Jhessabel Quino MenesesAún no hay calificaciones
- Guia Fisiopatologia 2017 IIDocumento8 páginasGuia Fisiopatologia 2017 IIUwOAún no hay calificaciones
- Historia de FarmaciaDocumento9 páginasHistoria de FarmaciaJackelin Jhessabel Quino MenesesAún no hay calificaciones
- Caso Cliníco DengueDocumento4 páginasCaso Cliníco DengueJackelin Jhessabel Quino MenesesAún no hay calificaciones
- Adriannn Maravi Exposicion de GastronomiaDocumento2 páginasAdriannn Maravi Exposicion de GastronomiaJackelin Jhessabel Quino MenesesAún no hay calificaciones
- Liderago Proyecto de VidaDocumento1 páginaLiderago Proyecto de VidaJackelin Jhessabel Quino MenesesAún no hay calificaciones
- Guía de Prácticas - Farmacobotánica - 1164596803Documento39 páginasGuía de Prácticas - Farmacobotánica - 1164596803Jackelin Jhessabel Quino MenesesAún no hay calificaciones
- TERMOSDocumento63 páginasTERMOSJackelin Jhessabel Quino MenesesAún no hay calificaciones
- Actividad Pagina 14 QuinoDocumento2 páginasActividad Pagina 14 QuinoJackelin Jhessabel Quino MenesesAún no hay calificaciones
- Rúbrica Semana 14-1Documento1 páginaRúbrica Semana 14-1Jackelin Jhessabel Quino MenesesAún no hay calificaciones
- Producto Final MatematicaDocumento2 páginasProducto Final MatematicaJackelin Jhessabel Quino MenesesAún no hay calificaciones
- Actividad 1 LiderazgoDocumento2 páginasActividad 1 LiderazgoJackelin Jhessabel Quino MenesesAún no hay calificaciones
- S10 Ejemplos Prueba de NormalidadDocumento12 páginasS10 Ejemplos Prueba de NormalidadJackelin Jhessabel Quino MenesesAún no hay calificaciones
- Semana 3 - Hidrogeno - Quino MenesesDocumento4 páginasSemana 3 - Hidrogeno - Quino MenesesJackelin Jhessabel Quino MenesesAún no hay calificaciones
- S03 Técnicas e Instrumentos y Matriz de DatosDocumento30 páginasS03 Técnicas e Instrumentos y Matriz de DatosJackelin Jhessabel Quino MenesesAún no hay calificaciones
- T06 QuiDocumento59 páginasT06 Quiapi-3711607100% (1)
- S03 Base Datos Credisa PhilcoDocumento6 páginasS03 Base Datos Credisa PhilcoJackelin Jhessabel Quino MenesesAún no hay calificaciones
- Programa Negociación Sandler TrainingDocumento2 páginasPrograma Negociación Sandler TrainingEdwin31OSRAún no hay calificaciones
- Manifiesto, El CreacionismoDocumento19 páginasManifiesto, El Creacionismojasan3000Aún no hay calificaciones
- TDHADocumento8 páginasTDHAfernanda davilaAún no hay calificaciones
- Rubrica-Ingles Vii HechoDocumento1 páginaRubrica-Ingles Vii HechoCrusvel Silva MedallaAún no hay calificaciones
- GPC, DHHQDocumento9 páginasGPC, DHHQRubi NuñezAún no hay calificaciones
- Agencia Metropolitana de TránsitoDocumento8 páginasAgencia Metropolitana de TránsitoKodi KodiAún no hay calificaciones
- Unidad 2: La Solidaridad y La Amistad: Evaluación Lengua y Literatura - Séptimo BásicoDocumento9 páginasUnidad 2: La Solidaridad y La Amistad: Evaluación Lengua y Literatura - Séptimo BásicoAlejandra VargasAún no hay calificaciones
- El Fraude en La Ciencia. El Caso HwangDocumento8 páginasEl Fraude en La Ciencia. El Caso HwangAndrea Marcela AguirreAún no hay calificaciones
- Los Parásitos y DesparasitantesDocumento61 páginasLos Parásitos y DesparasitantesAlejandro Aliendre100% (1)
- Marco Teórico-La Ventana de JohariDocumento2 páginasMarco Teórico-La Ventana de JohariMarvin Omar Caceres CanalesAún no hay calificaciones
- Fernando David Molina Jacome 6to Ciclo "C": SubjetivoDocumento5 páginasFernando David Molina Jacome 6to Ciclo "C": SubjetivoFernando David Molina JacomeAún no hay calificaciones
- Orientaciones para El Foro 2Documento3 páginasOrientaciones para El Foro 2jorge m sanchezAún no hay calificaciones
- Boletin Jesuita #73 Octubre 1981-Conducir en Lo Grande y en Lo PequeñoDocumento12 páginasBoletin Jesuita #73 Octubre 1981-Conducir en Lo Grande y en Lo PequeñoJuan Manuel Musante100% (1)
- Instrumentos Medievales en El Pórtico de La GloriaDocumento51 páginasInstrumentos Medievales en El Pórtico de La GloriaCarlos JofreAún no hay calificaciones
- Comunicaciòn Semana 2 (12 Al 16 de Abril)Documento5 páginasComunicaciòn Semana 2 (12 Al 16 de Abril)lidia yucra sullcapumaAún no hay calificaciones
- Situaciones Significativas CYT MARILÚDocumento2 páginasSituaciones Significativas CYT MARILÚMarilu PeraltaAún no hay calificaciones
- Exégesis Marcos 8, 22-26Documento2 páginasExégesis Marcos 8, 22-26Columbano80% (5)
- Asesinato en El Canadian ExpressDocumento10 páginasAsesinato en El Canadian ExpressLuis Andrés Martínez Figueroa67% (3)
- Números de BernoulliDocumento47 páginasNúmeros de BernoulliOscar GutierrezAún no hay calificaciones
- Investigación FormativaDocumento51 páginasInvestigación FormativaLesly PoloAún no hay calificaciones
- JOHNÇDocumento10 páginasJOHNÇBeauty In You 33Aún no hay calificaciones
- Factores Humanos PPLDocumento103 páginasFactores Humanos PPLcheesevakeAún no hay calificaciones
- Parcial 4 de Filosofia JuridicaDocumento7 páginasParcial 4 de Filosofia JuridicaMery Gomez Ferreyra100% (2)
- Introducción y ConclusionDocumento2 páginasIntroducción y ConclusionAldo AdriánAún no hay calificaciones
- Entrenar A Un San BernardoDocumento2 páginasEntrenar A Un San BernardoobedientdogAún no hay calificaciones
- Biomecanica de La Marcha Humana en Niño de 10 AñosDocumento23 páginasBiomecanica de La Marcha Humana en Niño de 10 AñosLisbeth ValenciaAún no hay calificaciones
- Tema Que Pasa Con Mi Cuerpo Cuando Me Muevo 09-06-2021Documento2 páginasTema Que Pasa Con Mi Cuerpo Cuando Me Muevo 09-06-2021nico100% (1)
- 223 - Kruse Martí Luter en Perspectiva EcumènicaDocumento11 páginas223 - Kruse Martí Luter en Perspectiva EcumènicaenriquearqueroAún no hay calificaciones
- Teoria 13Documento17 páginasTeoria 13Cristhian JesusAún no hay calificaciones
- Teoria de La PenaDocumento22 páginasTeoria de La PenaAnnySantaellaAún no hay calificaciones