También podría gustarte
- Resumen de La Búsqueda de Nuestros Ancestros: RESÚMENES UNIVERSITARIOSDe EverandResumen de La Búsqueda de Nuestros Ancestros: RESÚMENES UNIVERSITARIOSAún no hay calificaciones
- Un viaje por la prehistoriaDe EverandUn viaje por la prehistoriaCalificación: 4 de 5 estrellas4/5 (1)
- División de La Prehistoria y La HistoriaDocumento42 páginasDivisión de La Prehistoria y La Historiabetoking100% (3)
- Últimas noticias de sapiens: Revolución en nuestros orígenesDe EverandÚltimas noticias de sapiens: Revolución en nuestros orígenesAún no hay calificaciones
- Origen Del HombreDocumento18 páginasOrigen Del HombreJose Eduardo Arvayo Ovalles100% (1)
- AustralopithecusDocumento9 páginasAustralopithecusendersandovalAún no hay calificaciones
- Yacimientos Prehistóricos Del MundoDocumento32 páginasYacimientos Prehistóricos Del Mundobernarpi100% (3)
- Cuestionario HominizacionDocumento7 páginasCuestionario Hominizacionrenec_3Aún no hay calificaciones
- La Evolución Humana InestigacionDocumento4 páginasLa Evolución Humana InestigacionMarieling Soza100% (1)
- 03 - Origen Del Hombre (II) 2019-IIDocumento17 páginas03 - Origen Del Hombre (II) 2019-IIJhon Deyvi Sanchez TamayAún no hay calificaciones
- LA SALIDA DE ÁFRICA - MetodeDocumento7 páginasLA SALIDA DE ÁFRICA - MetodeCamila Mejía HenaoAún no hay calificaciones
- Evolución HumanaDocumento21 páginasEvolución Humanamayerly gonzalez cantilloAún no hay calificaciones
- Filogenia HumanaDocumento12 páginasFilogenia HumanaLuis Antonio Castillo RojasAún no hay calificaciones
- Resumen de Los Orígenes del Hombre de Richard Leakey: RESÚMENES UNIVERSITARIOSDe EverandResumen de Los Orígenes del Hombre de Richard Leakey: RESÚMENES UNIVERSITARIOSCalificación: 1 de 5 estrellas1/5 (1)
- Prehistoria I ApuntesDocumento53 páginasPrehistoria I ApuntesTeresa Jimenez HerreraAún no hay calificaciones
- T5 - Evolución y HominizaciónDocumento13 páginasT5 - Evolución y HominizaciónBel NavarroAún no hay calificaciones
- HomínidosDocumento1 páginaHomínidosblasito78100% (1)
- Lenguaje y Comunicación 2012 IDocumento4 páginasLenguaje y Comunicación 2012 IEfraín Wilmer Huamán ApolinarioAún no hay calificaciones
- Agustí - La Gran Migración U2Documento10 páginasAgustí - La Gran Migración U2francobrusatti2002Aún no hay calificaciones
- Resumen PemexDocumento209 páginasResumen PemexDahlia NieblaAún no hay calificaciones
- La Evolucion Del Ser HumanoDocumento10 páginasLa Evolucion Del Ser HumanoEduardo Geovanny Gomez CastroAún no hay calificaciones
- Etapas en La Evolución HumanaDocumento36 páginasEtapas en La Evolución HumanaJoansito Avila100% (1)
- Depredadores Hominidos y HumanosDocumento17 páginasDepredadores Hominidos y HumanosDaniel Sanchez100% (1)
- Hominidos Antes Del Homo SapiensDocumento18 páginasHominidos Antes Del Homo SapiensMaria Alejandra Hernandez RamirezAún no hay calificaciones
- AfropitecinosDocumento2 páginasAfropitecinosKEVIN JTAún no hay calificaciones
- Restos FósilesDocumento13 páginasRestos FósilesMónica AlvarezAún no hay calificaciones
- Linea de Tiempo HumanaDocumento34 páginasLinea de Tiempo HumanaFravioAún no hay calificaciones
- Evolucion Del HombreDocumento26 páginasEvolucion Del HombrecondexituAún no hay calificaciones
- ANTROPOGENESISDocumento8 páginasANTROPOGENESIShillins100% (1)
- Etapas en La Línea Evolutiva HumanaDocumento44 páginasEtapas en La Línea Evolutiva HumanaSilvy Gc50% (2)
- Ardipithecus RamidusDocumento24 páginasArdipithecus RamidusEmily MenaAún no hay calificaciones
- Nuestra Historia EvolutivaDocumento40 páginasNuestra Historia EvolutivaVcarrillo989100% (2)
- El Camino Evolutivo Del Hombre1Documento60 páginasEl Camino Evolutivo Del Hombre1chris meironeAún no hay calificaciones
- Australopithecus INTRODUCCIÓNDocumento7 páginasAustralopithecus INTRODUCCIÓNVictor DugarteAún no hay calificaciones
- Primates Fósiles 2Documento26 páginasPrimates Fósiles 2Cami BattistelliAún no hay calificaciones
- Taller 6 Evolucion HumanaDocumento4 páginasTaller 6 Evolucion HumanaAlejandro DelgadilloAún no hay calificaciones
- La HominizacionDocumento8 páginasLa HominizacionRrow MeisterAún no hay calificaciones
- 05 EVOLUCIÓN HUMANA - de Los Primates Al Homo Sapiens)Documento14 páginas05 EVOLUCIÓN HUMANA - de Los Primates Al Homo Sapiens)Ricardo ChavezAún no hay calificaciones
- Acosta EYGH20Documento17 páginasAcosta EYGH20Alvaro AcostaAún no hay calificaciones
- Evolucion HumanaDocumento7 páginasEvolucion HumanaKEVIN JOSE CARVAJAL MENJIVARAún no hay calificaciones
- Evolución Del HombreDocumento7 páginasEvolución Del HombreEsteban ValenciaAún no hay calificaciones
- Tema 1 El Hombre y La Cultura InicialDocumento74 páginasTema 1 El Hombre y La Cultura InicialMilagros Flores RumayAún no hay calificaciones
- Evolución HumanaDocumento22 páginasEvolución HumanaAlvaro AguilarAún no hay calificaciones
- Evolución Del Hombre InvestigaciónDocumento2 páginasEvolución Del Hombre Investigaciónyoru niAún no hay calificaciones
- Homo ArcaicoDocumento6 páginasHomo ArcaicoNicolas SerranoAún no hay calificaciones
- Etapas en La Evolución HumanaDocumento6 páginasEtapas en La Evolución Humanaduenmosh_kj3073100% (1)
- 6.evolucion de Hominidos-CorregidaDocumento33 páginas6.evolucion de Hominidos-CorregidaJoahan soto ortizAún no hay calificaciones
- Evolución de Los HomínidosDocumento46 páginasEvolución de Los Homínidosaltruista54Aún no hay calificaciones
- 274 VanderMade2014 GrandesMamiferosPlioPleistAfrica CunaHumanidadDocumento26 páginas274 VanderMade2014 GrandesMamiferosPlioPleistAfrica CunaHumanidadMarcelino BertranAún no hay calificaciones
- AustralopitecinosDocumento5 páginasAustralopitecinoselsuscrackAún no hay calificaciones
- 01-El Origen Del HombreDocumento27 páginas01-El Origen Del HombreVictor CisnerosAún no hay calificaciones
- Evolución HumanaDocumento72 páginasEvolución HumanaJhon RuizAún no hay calificaciones
- Evolución Del HombreDocumento3 páginasEvolución Del HombreJorge MartinezAún no hay calificaciones
- CUESTIONARIO LA HOMINIZACION Denisse CabreraDocumento7 páginasCUESTIONARIO LA HOMINIZACION Denisse CabreraDenisse Cabrera ChalasAún no hay calificaciones
- Africa Hominidos y El Origen Del HombreDocumento11 páginasAfrica Hominidos y El Origen Del Hombreleonel cAún no hay calificaciones
- Teorìa AntropogenèticaDocumento35 páginasTeorìa Antropogenèticaaguileraalejandro0% (1)
- Origen y Evolución de La HumanidadDocumento8 páginasOrigen y Evolución de La Humanidadjulenran1Aún no hay calificaciones
- Definición DeprehistoriaDocumento8 páginasDefinición DeprehistoriaRubriaJimenezAún no hay calificaciones
- Antecesores Del HombreDocumento7 páginasAntecesores Del HombreJaime Abregu67% (3)
- Evolucion Humana 2016 ERS1Documento79 páginasEvolucion Humana 2016 ERS1denisse antonia canales sotoAún no hay calificaciones
- Peña Hugo Deberes de HistoriaDocumento5 páginasPeña Hugo Deberes de HistoriaHugo Peña RamiroAún no hay calificaciones
- Diario de Clase Seminario 1 Hsce 2022-2023Documento3 páginasDiario de Clase Seminario 1 Hsce 2022-2023Hugo Peña RamiroAún no hay calificaciones
- La Musa Aprende A EscribirDocumento26 páginasLa Musa Aprende A EscribirHugo Peña RamiroAún no hay calificaciones
- Historia Del Pensamiento Político. TemasDocumento36 páginasHistoria Del Pensamiento Político. TemasHugo Peña RamiroAún no hay calificaciones
- Apuntes HsceDocumento22 páginasApuntes HsceHugo Peña RamiroAún no hay calificaciones
- Trabajo TutoríaDocumento8 páginasTrabajo TutoríaHugo Peña RamiroAún no hay calificaciones
- Apuntes de Historia Social de La Cultura Escrita Temas Del 1 Al 4Documento22 páginasApuntes de Historia Social de La Cultura Escrita Temas Del 1 Al 4Hugo Peña RamiroAún no hay calificaciones
- Prehistoria de Las Sociedades AmerindiasToda La TeoriaDocumento42 páginasPrehistoria de Las Sociedades AmerindiasToda La TeoriaHugo Peña RamiroAún no hay calificaciones
- Textos Pensamiento Político 2022-2023Documento21 páginasTextos Pensamiento Político 2022-2023Hugo Peña RamiroAún no hay calificaciones
- Reino PlantaeDocumento20 páginasReino PlantaeSendy Pérez100% (1)
- Examen 2 Unidad Biologia Chavez Minaya FrankDocumento15 páginasExamen 2 Unidad Biologia Chavez Minaya FrankFrank anthony Chávez minayaAún no hay calificaciones
- ADN MitocondrialDocumento3 páginasADN MitocondrialMateo MéndezAún no hay calificaciones
- Qheshwa SimiDocumento14 páginasQheshwa SimiIrving MurielAún no hay calificaciones
- Anexo T8Documento1 páginaAnexo T8Yuli MSAún no hay calificaciones
- Simulacro 6 - Area ADocumento14 páginasSimulacro 6 - Area AMiguel ValverdeAún no hay calificaciones
- Características de La Célula AnimalDocumento2 páginasCaracterísticas de La Célula AnimalRicardo SosaAún no hay calificaciones
- Plan de Aula Ciencias Naturales Febrero 2019Documento4 páginasPlan de Aula Ciencias Naturales Febrero 2019MARITZA NIPIAún no hay calificaciones
- A2 T4 ComputaciónDocumento2 páginasA2 T4 ComputaciónArturo mtzAún no hay calificaciones
- Examen 1Documento8 páginasExamen 1Joseph RebazaAún no hay calificaciones
- Evaluación 1 EcosistemasDocumento4 páginasEvaluación 1 Ecosistemasprofe.rhenriquez.smAún no hay calificaciones
- NOM-011-SEMARNAT-1996 - Musgo, Heno y DoradillaDocumento6 páginasNOM-011-SEMARNAT-1996 - Musgo, Heno y Doradillasmricardo2445Aún no hay calificaciones
- Biología - Area A - Semana 07 - Con ClavesDocumento3 páginasBiología - Area A - Semana 07 - Con ClavesYosthyn Stalyn Lazaro AcuñaAún no hay calificaciones
- Grupo C Examen Final 15 02 333154 Downloable 2520968Documento9 páginasGrupo C Examen Final 15 02 333154 Downloable 2520968Agustin FernandezAún no hay calificaciones
- Martinez, Lesslie y Patiño, Lizbeth PDFDocumento5 páginasMartinez, Lesslie y Patiño, Lizbeth PDFLiz Patiño HuanéAún no hay calificaciones
- Genetica CompletoDocumento77 páginasGenetica CompletomarianoramonAún no hay calificaciones
- Anatomia de La Mano DorsalDocumento24 páginasAnatomia de La Mano DorsalCARLOSAún no hay calificaciones
- El Cóndor AndinoDocumento12 páginasEl Cóndor AndinoAlejandro Miranda DíazAún no hay calificaciones
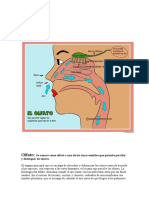
- Sentido Del OlfatoDocumento3 páginasSentido Del OlfatoJavier Escobar100% (1)
- TFG Jun20 Rodriguez Basarte PaulaDocumento69 páginasTFG Jun20 Rodriguez Basarte PaulaEnrique Ramos RodriguezAún no hay calificaciones
- Preguntas IcfesDocumento31 páginasPreguntas IcfesSandra Yulieth Santiago VegaAún no hay calificaciones
- Preguntas Unidad 1 y 2Documento17 páginasPreguntas Unidad 1 y 2Freddy SantiagoAún no hay calificaciones
- Histología CelularDocumento43 páginasHistología CelularXavier SaiwatabaAún no hay calificaciones
- Semana Del 12 Al 17 de DiciembreDocumento104 páginasSemana Del 12 Al 17 de DiciembreDanya LopezAún no hay calificaciones
- Anatomía y Fisiología Del Globo Ocular.Documento4 páginasAnatomía y Fisiología Del Globo Ocular.VIANNEY SANTOS MORALESAún no hay calificaciones
- Tema 2 Regla Del Tanto Por CientoDocumento1 páginaTema 2 Regla Del Tanto Por CientojjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjAún no hay calificaciones
- Olalla-Robert Louis StevensonDocumento108 páginasOlalla-Robert Louis StevensonCamAún no hay calificaciones
- Caracterización Anatómica de La Madera y Carbón de Cinco Especies Leñosas para Su IdentificacionDocumento19 páginasCaracterización Anatómica de La Madera y Carbón de Cinco Especies Leñosas para Su IdentificacionWilliam Aguilera Hancco100% (1)
- Diferencias Entre Dineina y KinesinaDocumento9 páginasDiferencias Entre Dineina y KinesinaJesenia Jeanet PHAún no hay calificaciones
- Hemostasia y Coagulación SanguíneaDocumento13 páginasHemostasia y Coagulación SanguíneaAngii LópezAún no hay calificaciones