También podría gustarte
- Resumen-Inside JobDocumento4 páginasResumen-Inside JobKristi Escobar80% (5)
- Test para Descubrir El Temperamento Segun HipocratesDocumento7 páginasTest para Descubrir El Temperamento Segun HipocratesKimy Pierina Pezo FrancoAún no hay calificaciones
- Matemáticas financieras UJAT e-book caracteresDocumento209 páginasMatemáticas financieras UJAT e-book caracteresAna Lilia Valle Santos27% (11)
- Los Ríos ProfundosDocumento6 páginasLos Ríos ProfundosEduar StivenAún no hay calificaciones
- ANTIESTRESDocumento6 páginasANTIESTRESMelisa Cruz Jaramillo100% (1)
- Mentiras OficialesDocumento31 páginasMentiras OficialespatrisainzAún no hay calificaciones
- Como Dar Glamour A Tu Marca Personal-Ana Jiménez PDFDocumento161 páginasComo Dar Glamour A Tu Marca Personal-Ana Jiménez PDFAdrianRodriguezAún no hay calificaciones
- Banca Comercial InternacionalDocumento4 páginasBanca Comercial InternacionalOriana Sánchez100% (2)
- Evolución de la vida a través de las eras geológicasDocumento2 páginasEvolución de la vida a través de las eras geológicasjhomar4559871% (7)
- INVESTIGACIÓN Y CONSERVACIÓN SOBRE MURCIÉLAGOS EN EL ECUADORDe EverandINVESTIGACIÓN Y CONSERVACIÓN SOBRE MURCIÉLAGOS EN EL ECUADORAún no hay calificaciones
- Primer Examen SimulacroDocumento7 páginasPrimer Examen SimulacroAngeles ReyesAún no hay calificaciones
- Taller3-Biogeografia-DerivaContinetal 2023Documento17 páginasTaller3-Biogeografia-DerivaContinetal 2023sancheza21Aún no hay calificaciones
- Stucchi Et Al 2009 - Oso Andino (Tremarctos Ornatus) Fósil de Chaquil - AmazonasDocumento6 páginasStucchi Et Al 2009 - Oso Andino (Tremarctos Ornatus) Fósil de Chaquil - AmazonasAICBPERUAún no hay calificaciones
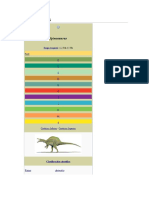
- Spinosaurus - Wikipedia, La Enciclopedia LibreDocumento81 páginasSpinosaurus - Wikipedia, La Enciclopedia LibreGerman Montes De OcaAún no hay calificaciones
- Wealden - Armoured - Dinosaurs (1) .En - EsDocumento30 páginasWealden - Armoured - Dinosaurs (1) .En - EsIgnacio NavaAún no hay calificaciones
- Evolucion y Filogenia de Arthropoda 500Documento10 páginasEvolucion y Filogenia de Arthropoda 500Maddy ChanAún no hay calificaciones
- OrigEvoDomMamifChileDocumento17 páginasOrigEvoDomMamifChileayaleytonAún no hay calificaciones
- El Gran Intercambio Biótico AmericanoDocumento21 páginasEl Gran Intercambio Biótico AmericanoCarina BugliaAún no hay calificaciones
- Spinosaurus (Wikipedia)Documento10 páginasSpinosaurus (Wikipedia)EnzoAún no hay calificaciones
- El Gran Intercambio Biótico AmericanoDocumento19 páginasEl Gran Intercambio Biótico AmericanoPablo Adrián OteroAún no hay calificaciones
- Evolución y Paleobiogeografía de Los Monos Del Nuevo MundoDocumento4 páginasEvolución y Paleobiogeografía de Los Monos Del Nuevo MundoJulian Santiago Casallas OsorioAún no hay calificaciones
- Tea de Paleo Final para Samir FINALLLDocumento31 páginasTea de Paleo Final para Samir FINALLLBetzy Valery Poma PerezAún no hay calificaciones
- SpinosaurusDocumento13 páginasSpinosaurusAzter KevinAún no hay calificaciones
- Arcosaurios No Avianos de ChileDocumento56 páginasArcosaurios No Avianos de ChilefrancoAún no hay calificaciones
- Paleobiogeografia y Paleoecologia de Los Mamiferos Continentales de America Del Sur Durante El Cretacico Tardio PaleocenoDocumento12 páginasPaleobiogeografia y Paleoecologia de Los Mamiferos Continentales de America Del Sur Durante El Cretacico Tardio PaleocenoGonzalo Galvez CardenasAún no hay calificaciones
- Una Nueva Especie de Neuryurus AmeghinoDocumento11 páginasUna Nueva Especie de Neuryurus Ameghinoeduardo ghioAún no hay calificaciones
- Presentacion ExposicionDocumento35 páginasPresentacion ExposicionManuel otayaAún no hay calificaciones
- Dossier Hominidos I 2022-2023Documento29 páginasDossier Hominidos I 2022-2023Hugo Peña RamiroAún no hay calificaciones
- Solucion Actividad 12Documento3 páginasSolucion Actividad 12Valeria TapiaAún no hay calificaciones
- Tercer Capitulo Entomologia - En.esDocumento53 páginasTercer Capitulo Entomologia - En.esBrianm FlorezAún no hay calificaciones
- Chornogubsky y Kramarz. 2012 - Microbiotheriidae FM PinturasDocumento9 páginasChornogubsky y Kramarz. 2012 - Microbiotheriidae FM PinturasLauraAún no hay calificaciones
- CarcharodontosaurusDocumento46 páginasCarcharodontosaurusSaul de la VegaAún no hay calificaciones
- Origen de La Biota Antillana Contada Por Amblipígidos (Arachnida Amblypygi) - Jose CruzDocumento13 páginasOrigen de La Biota Antillana Contada Por Amblipígidos (Arachnida Amblypygi) - Jose CruzJose Armando Cruz LimaAún no hay calificaciones
- Documento Completo - PDF PDFADocumento12 páginasDocumento Completo - PDF PDFAJosue VelasquezAún no hay calificaciones
- Cretácico en ArgentinaDocumento17 páginasCretácico en Argentinapablo314Aún no hay calificaciones
- SpinosaurusDocumento21 páginasSpinosaurusPedrito VFAún no hay calificaciones
- Ensayo Origen y Evolucion ArtropodosDocumento8 páginasEnsayo Origen y Evolucion ArtropodosIvan Rivera100% (1)
- Texto ValentinaDocumento2 páginasTexto ValentinaJaviera ArriazaAún no hay calificaciones
- Geología de Cuba - EcuRedDocumento3 páginasGeología de Cuba - EcuRedVictoriano NietoAún no hay calificaciones
- Álbum de Biología Belén Guevara11moDocumento13 páginasÁlbum de Biología Belén Guevara11moBelén GuevaraAún no hay calificaciones
- Especies de HomínidosDocumento11 páginasEspecies de Homínidosnaturaleseem1de5Aún no hay calificaciones
- 6.evolucion de Hominidos-CorregidaDocumento33 páginas6.evolucion de Hominidos-CorregidaJoahan soto ortizAún no hay calificaciones
- Descripción General: Triceratops Megalosaurus IguanodonDocumento3 páginasDescripción General: Triceratops Megalosaurus IguanodonEmiLuz Q MendozaAún no hay calificaciones
- TejadaEtAl2009b Fósiles de Espinar, Cusco PDFDocumento6 páginasTejadaEtAl2009b Fósiles de Espinar, Cusco PDFantony huillca bornasAún no hay calificaciones
- Megalosaurus descripción terópodo JurásicoDocumento3 páginasMegalosaurus descripción terópodo JurásicoJhean YgnacioAún no hay calificaciones
- Australopithecus AfricanusDocumento11 páginasAustralopithecus AfricanusNoelia CaligariAún no hay calificaciones
- Proyecto de InvestigaciónDocumento9 páginasProyecto de InvestigaciónHuascar MejiaAún no hay calificaciones
- Origen y Evolución de La HumanidadDocumento8 páginasOrigen y Evolución de La Humanidadjulenran1Aún no hay calificaciones
- Registro fósil OaxacaDocumento7 páginasRegistro fósil OaxacaLeslie LouAún no hay calificaciones
- Aves Del TerrorDocumento8 páginasAves Del Terrorcontreras.varela.zuleyma.jAún no hay calificaciones
- Biología Zarigüeya ComúnDocumento13 páginasBiología Zarigüeya ComúnLolelo1234567890Aún no hay calificaciones
- WG - Dinosaurios - v12.08Documento3729 páginasWG - Dinosaurios - v12.08smodkaAún no hay calificaciones
- Plantilla Del Texto ExpositivoDocumento6 páginasPlantilla Del Texto ExpositivoKarem GómezAún no hay calificaciones
- Royo-TorresR 2009dinosau Sauropodospen IbricaActasIVjornadasintern SalasInfantes 2007Documento29 páginasRoyo-TorresR 2009dinosau Sauropodospen IbricaActasIVjornadasintern SalasInfantes 2007Naiara Dominguez GilAún no hay calificaciones
- ToumaiDocumento2 páginasToumaiAlfaAún no hay calificaciones
- Artículo de Revisión GliptodonteDocumento6 páginasArtículo de Revisión GliptodonteLeidy FlórezAún no hay calificaciones
- XIII Congreso Geologico Antofagasta 2012 Paleontologia T6Documento49 páginasXIII Congreso Geologico Antofagasta 2012 Paleontologia T6dg_venegasAún no hay calificaciones
- Zeledon 2006. Notas Sobre Equinodermos Fósiles de Costa Rica AlvaradoDocumento14 páginasZeledon 2006. Notas Sobre Equinodermos Fósiles de Costa Rica AlvaradoelibrenAún no hay calificaciones
- Test y Repuestas Prehistoria I. C. R.C.Documento10 páginasTest y Repuestas Prehistoria I. C. R.C.alfredoAún no hay calificaciones
- 13.OrigEvoDomMamifChileDocumento18 páginas13.OrigEvoDomMamifChileclaudiaAún no hay calificaciones
- Zoologia de Invertebrados 2Documento33 páginasZoologia de Invertebrados 2AlejanDrah BarriiOsAún no hay calificaciones
- Australopithecus AfricanusDocumento11 páginasAustralopithecus AfricanusNoelia CaligariAún no hay calificaciones
- Fósiles Encontrados en La Ampliación Del Canal de PanamaDocumento25 páginasFósiles Encontrados en La Ampliación Del Canal de Panamayaneidys alfonsoAún no hay calificaciones
- Dinosaurios MersDocumento11 páginasDinosaurios Mersdavid cetinaAún no hay calificaciones
- Los Dinosaurios AmericanosDocumento4 páginasLos Dinosaurios AmericanosLourdes Landin XD75% (4)
- Carrilloetal2008 RegistromastodontenenVenezuelaDocumento32 páginasCarrilloetal2008 RegistromastodontenenVenezuelaJesus lozadaAún no hay calificaciones
- Diaz Etal2023 - Ksima LaGuaira - BIOLLANIA V16 - pp113 123Documento11 páginasDiaz Etal2023 - Ksima LaGuaira - BIOLLANIA V16 - pp113 123teddy ramirezAún no hay calificaciones
- Dinosaurios y Piedras de Ica: evidencia de coexistenciaDocumento23 páginasDinosaurios y Piedras de Ica: evidencia de coexistenciaHéctor rivadeneyraAún no hay calificaciones
- Resumen de La Búsqueda de Nuestros Ancestros: RESÚMENES UNIVERSITARIOSDe EverandResumen de La Búsqueda de Nuestros Ancestros: RESÚMENES UNIVERSITARIOSAún no hay calificaciones
- Prueba de Lectura Barcos Que Vuelan PruebaDocumento4 páginasPrueba de Lectura Barcos Que Vuelan PruebaRaul Iriarte AnayaAún no hay calificaciones
- Huaca Historia 1Documento10 páginasHuaca Historia 1Dani GuevaraAún no hay calificaciones
- 5 Experimentos de ClonaciónDocumento5 páginas5 Experimentos de ClonaciónRoger GomezAún no hay calificaciones
- Cronología de Las Funciones Del TRABAJADOR SOCIALDocumento17 páginasCronología de Las Funciones Del TRABAJADOR SOCIALALIZTRAVIEZAAún no hay calificaciones
- 07 Remuneración y Satisfacción LaboralDocumento17 páginas07 Remuneración y Satisfacción LaboralYesenia Cruz ParedesAún no hay calificaciones
- Presentacion Cap 12 PMI UPCDocumento79 páginasPresentacion Cap 12 PMI UPCguillevallejogAún no hay calificaciones
- Catecismo 478Documento4 páginasCatecismo 478dulceAún no hay calificaciones
- Globo Ocular o Bulbo Del OjoDocumento14 páginasGlobo Ocular o Bulbo Del OjoFreddy PozoAún no hay calificaciones
- Guia Juan CastroDocumento3 páginasGuia Juan Castrolizkatherine198150% (2)
- Listado Transacciones SaP HCMDocumento11 páginasListado Transacciones SaP HCMEstrellitaHdzHdzAún no hay calificaciones
- Privado VIII (Daños) Resumen de Bustamante AlsinaDocumento84 páginasPrivado VIII (Daños) Resumen de Bustamante AlsinamarinaAún no hay calificaciones
- Madre Coraje y Sus Hijos CriticaDocumento7 páginasMadre Coraje y Sus Hijos CriticaFlorencia PazAún no hay calificaciones
- MM 0701670300Documento2 páginasMM 0701670300joseAún no hay calificaciones
- Glovalizacion de La Arquitectura ContemporaneaDocumento10 páginasGlovalizacion de La Arquitectura ContemporaneavalerieAún no hay calificaciones
- Casos de Fracionamiento Parcial - 55920502Documento9 páginasCasos de Fracionamiento Parcial - 55920502NELSON ADOLFO DAZA HERRERAAún no hay calificaciones
- Guía de Ruta y Avance de Ruta para La Realimentación - Fase 3. Paz ColombiaDocumento5 páginasGuía de Ruta y Avance de Ruta para La Realimentación - Fase 3. Paz ColombiaSTEFAAún no hay calificaciones
- Unidad LV. Actividad LLL - Ramas Del Derecho Publico.Documento2 páginasUnidad LV. Actividad LLL - Ramas Del Derecho Publico.Carliset López BatistaAún no hay calificaciones
- Catalogo NGDocumento96 páginasCatalogo NGcva201717Aún no hay calificaciones
- Modelo para Administrar El Cambio Organizacional LEWINDocumento1 páginaModelo para Administrar El Cambio Organizacional LEWINEmilio CulajayAún no hay calificaciones
- 4 - Najmanovich, D. Estética de La ComplejidadDocumento30 páginas4 - Najmanovich, D. Estética de La ComplejidadMario Apolinar RuizAún no hay calificaciones
- Institución Universitaria Politécnico GrancolombianoDocumento12 páginasInstitución Universitaria Politécnico GrancolombianoAlejandra PinzónAún no hay calificaciones