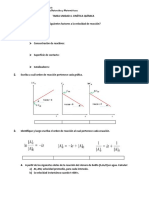
También podría gustarte
- Cinética EnzimáticaDocumento12 páginasCinética EnzimáticaMaria Alejandra SantanderAún no hay calificaciones
- Problemas de Recacciones de Orden y Vida Media de CineticaDocumento46 páginasProblemas de Recacciones de Orden y Vida Media de CineticaHugo AlemánAún no hay calificaciones
- Cinética QuímicaDocumento8 páginasCinética QuímicaGinger Damaris Patso LoorAún no hay calificaciones
- Practica 8 Yodación de AcetonaDocumento9 páginasPractica 8 Yodación de AcetonaKiiKi DeeAún no hay calificaciones
- Normativa 7Documento6 páginasNormativa 7paula100% (4)
- Tarea 01-Consolidado 1-Analisis Estructural IiDocumento20 páginasTarea 01-Consolidado 1-Analisis Estructural IiYZARRA PEREZ JOEL100% (1)
- BQ-I Tema 5Documento27 páginasBQ-I Tema 5Carmen GómezAún no hay calificaciones
- TP 1Documento12 páginasTP 1Aguss ColettaAún no hay calificaciones
- Sesiã N 14 - UV - Cinã©tica de Las ReaccionesDocumento39 páginasSesiã N 14 - UV - Cinã©tica de Las ReaccionesMiyaray LecarosAún no hay calificaciones
- Reacciones ComplejasDocumento26 páginasReacciones ComplejasErika Itzel C AAún no hay calificaciones
- Cinetica TallerDocumento10 páginasCinetica TallerStiven SofanAún no hay calificaciones
- Cinética Química - ClaseDocumento28 páginasCinética Química - Clasederekaquino2000Aún no hay calificaciones
- Cinética de La Hidrólisis Del Acetato de MetiloDocumento8 páginasCinética de La Hidrólisis Del Acetato de MetiloRichard Cáceres100% (1)
- Enz-3 - 2019 Enzimas-CineticaDocumento58 páginasEnz-3 - 2019 Enzimas-CineticaRomina Garavito-Salini CasasAún no hay calificaciones
- Laboratorio 4 - Quim. ExpDocumento7 páginasLaboratorio 4 - Quim. ExpDavid Alejandro RamirezAún no hay calificaciones
- Ejercicios Unidad 2 HidrometalurgiaDocumento9 páginasEjercicios Unidad 2 HidrometalurgiabiankajmzlAún no hay calificaciones
- Cinetica 2013Documento30 páginasCinetica 2013JuliAún no hay calificaciones
- Cinetica EnzimaticaDocumento23 páginasCinetica EnzimaticaMariana Maya GuerreroAún no hay calificaciones
- Cinética Química (Química 2º de Bachillerato)Documento56 páginasCinética Química (Química 2º de Bachillerato)frodríguez_30Aún no hay calificaciones
- Teoría Unidad 8Documento93 páginasTeoría Unidad 8Roger SalasAún no hay calificaciones
- Fisico Quimica (Cinética Química)Documento41 páginasFisico Quimica (Cinética Química)Barbara DumasAún no hay calificaciones
- Guia PRACTICA No 7Documento3 páginasGuia PRACTICA No 7yesikaAún no hay calificaciones
- Exposicion de Cinetica QuimicaDocumento48 páginasExposicion de Cinetica QuimicaJuan Villalobos Nuñez100% (1)
- Cinetica II 2Documento35 páginasCinetica II 2Patricia Emperatriz LopezaguirreAún no hay calificaciones
- 17 Cinetica Quimica Parte 1Documento40 páginas17 Cinetica Quimica Parte 1Jose BanchonAún no hay calificaciones
- CUESTIONARIODocumento11 páginasCUESTIONARIORicardo CortezAún no hay calificaciones
- Introdución Fotometría. Ley de BBDocumento30 páginasIntrodución Fotometría. Ley de BBSamuel DoomWyldeAún no hay calificaciones
- Cinética Química Ginger PatsoDocumento8 páginasCinética Química Ginger PatsoGinger Damaris Patso LoorAún no hay calificaciones
- Taller 2 - Reacciones 1Documento7 páginasTaller 2 - Reacciones 1Joel BasantesAún no hay calificaciones
- Semana 11 - San Marcos - Nov - 2019 PDFDocumento29 páginasSemana 11 - San Marcos - Nov - 2019 PDFYudScarpAún no hay calificaciones
- Problemas de Bioquímica - Primer Curso Del Grado en Bioquímica - Curso 2010 11Documento8 páginasProblemas de Bioquímica - Primer Curso Del Grado en Bioquímica - Curso 2010 11cristipisti88Aún no hay calificaciones
- Reactor de Mezcla CompletaDocumento11 páginasReactor de Mezcla CompletaClaudia GonzalesAún no hay calificaciones
- Evaluación 2 Biotecnología de Alimentos Caceres Quispe AlejandraDocumento12 páginasEvaluación 2 Biotecnología de Alimentos Caceres Quispe AlejandramcaceresquiAún no hay calificaciones
- Tercer Tutorial - Grupo 5Documento12 páginasTercer Tutorial - Grupo 5Amed Enrique Ortega MonteroAún no hay calificaciones
- Taller EnzimasDocumento5 páginasTaller Enzimaswilber menesesAún no hay calificaciones
- Ejercicios Resueltos - Reactores QuímicosDocumento5 páginasEjercicios Resueltos - Reactores QuímicosMABAún no hay calificaciones
- Fisicoquímica Atmosférica Fase 2 - AireDocumento14 páginasFisicoquímica Atmosférica Fase 2 - AiremariaAún no hay calificaciones
- Tarea Cinética Química AaDocumento4 páginasTarea Cinética Química Aagustavo valdiviesoAún no hay calificaciones
- Diseño ReacDocumento10 páginasDiseño ReacivanAún no hay calificaciones
- Tutorial 7Documento8 páginasTutorial 7anani quispeAún no hay calificaciones
- Taller de Cinética QuímicaDocumento13 páginasTaller de Cinética QuímicaClaudia GuerreroAún no hay calificaciones
- Practica 2 Lab-FQIII-1Documento11 páginasPractica 2 Lab-FQIII-1Alejandra ZarateAún no hay calificaciones
- Solucions Prob EnzimsDocumento26 páginasSolucions Prob EnzimsAlbert Crespo RamiroAún no hay calificaciones
- Práctica CalificadacineticaDocumento2 páginasPráctica CalificadacineticaLuis APAún no hay calificaciones
- Cinetica EnzimaticaDocumento21 páginasCinetica EnzimaticapotatochipAún no hay calificaciones
- 02 CinéticaDocumento55 páginas02 CinéticayunmymendozagarciaAún no hay calificaciones
- Cinética QuímicaDocumento58 páginasCinética QuímicaEDUARDO SAMIR ROMAN VIVASAún no hay calificaciones
- Unidad 3 1. Cinética Generalidades Conceptos BásicosDocumento29 páginasUnidad 3 1. Cinética Generalidades Conceptos BásicosJoshua PazmiñoAún no hay calificaciones
- G9 - Cinética - 27 - 7 - 20Documento6 páginasG9 - Cinética - 27 - 7 - 20josAún no hay calificaciones
- Biotecnología Guía PrácticosDocumento13 páginasBiotecnología Guía PrácticosPaula MartoglioAún no hay calificaciones
- Actividaad de TermoDocumento8 páginasActividaad de TermoKim TêteAún no hay calificaciones
- Clase 6 - 07-09-2022Documento10 páginasClase 6 - 07-09-2022Lucas RFAún no hay calificaciones
- TP5 MecanicaDocumento15 páginasTP5 Mecanicadavidgyt12Aún no hay calificaciones
- Taller BioquimicaDocumento10 páginasTaller BioquimicaAna María Muñoz JaramilloAún no hay calificaciones
- 02 CinéticaDocumento64 páginas02 CinéticaJavier MartinezAún no hay calificaciones
- Solucionario Del Examen Final de Fisicoquímica Ii 2022-Ii - MpucaDocumento6 páginasSolucionario Del Examen Final de Fisicoquímica Ii 2022-Ii - MpucaFABRICIO TELMO MATIAS MALCA SANCHEZAún no hay calificaciones
- CQCL R5Documento10 páginasCQCL R5DanielMoraAún no hay calificaciones
- Balance de Masa o MateriaDocumento16 páginasBalance de Masa o MateriaMauricio Ignacio García MoralesAún no hay calificaciones
- Clase 4 Reacciones y Cinética FinalDocumento37 páginasClase 4 Reacciones y Cinética FinalRosa Dina Gaona0% (1)
- Cinetica EnzimaticaDocumento40 páginasCinetica EnzimaticaJaime Polo100% (1)
- Ejercicio 2Documento2 páginasEjercicio 2jeisson santamariaAún no hay calificaciones
- La Fiesta Del Burrito SomoteñoDocumento3 páginasLa Fiesta Del Burrito SomoteñoOmán David DíazAún no hay calificaciones
- Guía 3 Origen y Composición Del Universo - 6Documento2 páginasGuía 3 Origen y Composición Del Universo - 6Jose Ricardo Riaño BarretoAún no hay calificaciones
- Materia SecaDocumento12 páginasMateria SecaIsmael Alessandro Peña CoronadoAún no hay calificaciones
- Valores Pueblos AndinosDocumento4 páginasValores Pueblos Andinosyury ramosAún no hay calificaciones
- MovadoFY17 SPDocumento70 páginasMovadoFY17 SPLuis David Morales cameyAún no hay calificaciones
- Infiltraciones en El PieDocumento10 páginasInfiltraciones en El PieIroNy100% (1)
- TODODocumento84 páginasTODOFabian MiñoAún no hay calificaciones
- Examen InglessDocumento1 páginaExamen InglessBetsabe MalpicaAún no hay calificaciones
- Informe YodometriaDocumento3 páginasInforme YodometriaISABELA RENTERIA RUIZAún no hay calificaciones
- Partes y Exigencias de Cultivo de La Caña de AzucarDocumento3 páginasPartes y Exigencias de Cultivo de La Caña de AzucarBrayan Chavez OréAún no hay calificaciones
- Culto A La PacchamamaDocumento6 páginasCulto A La PacchamamaMamerto Huaman AyteAún no hay calificaciones
- Informe Individualizado de Recuperación. 2º Bachillerato. GeografíaDocumento5 páginasInforme Individualizado de Recuperación. 2º Bachillerato. GeografíaArGeoHistoAún no hay calificaciones
- Flujo de Caja Cut and FillDocumento96 páginasFlujo de Caja Cut and FillOscar Esquivel TraslaviñaAún no hay calificaciones
- Hongos Comestibles (Programa)Documento5 páginasHongos Comestibles (Programa)Micaela GattiAún no hay calificaciones
- Manual de Inseminación Artificial 2Documento27 páginasManual de Inseminación Artificial 2Diego Fernando Vega ChimarroAún no hay calificaciones
- Biografia de Arne Naes1Documento6 páginasBiografia de Arne Naes1Valeria Vasquez PorrasAún no hay calificaciones
- Manufactura CelularDocumento8 páginasManufactura CelularLalo PadillaAún no hay calificaciones
- Practica 2 Vias de AdministracionDocumento13 páginasPractica 2 Vias de AdministracionNaomi Alcántara AmigónAún no hay calificaciones
- Derechos de La Mujer PoliciaDocumento171 páginasDerechos de La Mujer PoliciaWILLIAM LOPEZ RAMIREZAún no hay calificaciones
- CIRCULAR INFORMATIVA No.28Documento1 páginaCIRCULAR INFORMATIVA No.28rubenshoAún no hay calificaciones
- Resumen Fundamentos Legales de La Salud Ocupacional en Colombia.Documento35 páginasResumen Fundamentos Legales de La Salud Ocupacional en Colombia.TatianaAún no hay calificaciones
- Lab 7Documento9 páginasLab 7edison ccañiAún no hay calificaciones
- SEPTIMO Modulo SistemasDocumento35 páginasSEPTIMO Modulo SistemasYuli BetancurAún no hay calificaciones
- Crónica de Una Muerte Anunciada. Los Límites Del CrecimientoDocumento4 páginasCrónica de Una Muerte Anunciada. Los Límites Del Crecimientochavezlauri1100% (1)
- Influencia de La Arquitectura Hospitalaria en El Mejoramiento DelDocumento7 páginasInfluencia de La Arquitectura Hospitalaria en El Mejoramiento DelESTEBAN PATRICIO HIDALGO JARAMILLOAún no hay calificaciones
- Catalogo Sarel ElecinDocumento6 páginasCatalogo Sarel Elecinpier_89_9100% (1)
- Manual Vol 2Documento18 páginasManual Vol 2Montserrat HernándezAún no hay calificaciones
- Espiritualidad Cósmica AndinaDocumento4 páginasEspiritualidad Cósmica AndinaNico LMAún no hay calificaciones