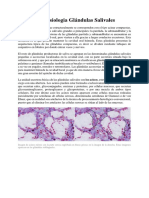
También podría gustarte
- Glandulas SalivalesDocumento9 páginasGlandulas SalivalesSamari BautistaAún no hay calificaciones
- Adenómeros GlandularesDocumento2 páginasAdenómeros GlandularesEdward A. De Los SantosAún no hay calificaciones
- Glándulas SalivalesDocumento7 páginasGlándulas SalivalesSamantha Heinrichs DomínguezAún no hay calificaciones
- Histología Glándulas SalivalesDocumento5 páginasHistología Glándulas SalivalesGROVER ALVARO POMA APAZAAún no hay calificaciones
- Estructura Histologica General de Las Glandulas SalivalesDocumento10 páginasEstructura Histologica General de Las Glandulas SalivalesSamari BautistaAún no hay calificaciones
- Preguntas Glndulas SalivalesDocumento9 páginasPreguntas Glndulas SalivalesSebastián Estay SaavedraAún no hay calificaciones
- Funciones de La Mucosa DigestivaDocumento4 páginasFunciones de La Mucosa Digestivajulio cesar oteroAún no hay calificaciones
- Capitulo 8 EmbriohistoDocumento37 páginasCapitulo 8 EmbriohistoKathya DiazAún no hay calificaciones
- Apuntes de Glandulas SalivalesDocumento11 páginasApuntes de Glandulas SalivalesTamara CárdenasAún no hay calificaciones
- Histofisiologia Glándulas SalivalesDocumento4 páginasHistofisiologia Glándulas SalivalesAlexiaBowenHerasAún no hay calificaciones
- Anatomía Glandulas Del CaninoDocumento12 páginasAnatomía Glandulas Del CaninoJorddan Andres Intriago MaciasAún no hay calificaciones
- Cap.8 Glándulas Salivales Embriohistología IIDocumento41 páginasCap.8 Glándulas Salivales Embriohistología IIPaolaAún no hay calificaciones
- Glandulas Anexas Del Tubo DigestivoDocumento5 páginasGlandulas Anexas Del Tubo DigestivoKaterine Lisbeth Carpio QuichcaAún no hay calificaciones
- Exposicion de Glandulas Salivales y TonsilaresDocumento7 páginasExposicion de Glandulas Salivales y TonsilaresJosselyn LoorAún no hay calificaciones
- Resumen Glandulas Salivares Resumen 1-BDocumento51 páginasResumen Glandulas Salivares Resumen 1-BIsabel FeblesAún no hay calificaciones
- Glandulas SalivalesDocumento13 páginasGlandulas SalivalesChristopher Gallo TapiaAún no hay calificaciones
- Digestivo PatoDocumento16 páginasDigestivo PatoMaria Jose Padilla Guilloth0% (1)
- (EV) S.digestivo III, GlandulasDocumento10 páginas(EV) S.digestivo III, GlandulasareliAún no hay calificaciones
- Actividad 1 Visualización de Las Glándulas SalivalesDocumento7 páginasActividad 1 Visualización de Las Glándulas Salivalespatricia mata moyaAún no hay calificaciones
- Las Glandulas SalivalesDocumento2 páginasLas Glandulas SalivalesGabriela TejadaAún no hay calificaciones
- Caracteristicas Estructurales Del HepatocitoDocumento7 páginasCaracteristicas Estructurales Del HepatocitoFernando Ibañez100% (2)
- Glándulas SalivalesDocumento95 páginasGlándulas SalivalesNaikaryAún no hay calificaciones
- Clase Semana #15 - Dr. RequenaDocumento2 páginasClase Semana #15 - Dr. RequenaKevin García MurgaAún no hay calificaciones
- Glándulas SalivalesDocumento24 páginasGlándulas SalivalesMarisol VargasAún no hay calificaciones
- GLÁNDULASDocumento122 páginasGLÁNDULASShota KunAún no hay calificaciones
- Glándulas SalivalesDocumento17 páginasGlándulas SalivalesMaria de Jesus100% (1)
- Epitelio Cubico EstratificadoDocumento11 páginasEpitelio Cubico EstratificadoDomenica LopezAún no hay calificaciones
- Histologia de Las Glandulas SalivalesDocumento9 páginasHistologia de Las Glandulas SalivalesEdmar Angel Aguilar LeyvaAún no hay calificaciones
- Glandulas SalivalesDocumento15 páginasGlandulas SalivalesAlejandroValenciaAún no hay calificaciones
- Tejido Epitelial GlandularDocumento8 páginasTejido Epitelial GlandularJudith Capel OrtegaAún no hay calificaciones
- Histologia Grupo 6..Documento15 páginasHistologia Grupo 6..MabeAún no hay calificaciones
- 3 Histología de Los SentidosDocumento57 páginas3 Histología de Los SentidosValery Ornella Timana EspinozaAún no hay calificaciones
- Sistema Digestivo-Glandulas AnexasDocumento41 páginasSistema Digestivo-Glandulas AnexasEnrique AranaAún no hay calificaciones
- Lengua y Glandulas SalivalesDocumento7 páginasLengua y Glandulas SalivalesMaria Noel Acuña MoralesAún no hay calificaciones
- Glandulas Anexas Al Sistema Digestivo.Documento25 páginasGlandulas Anexas Al Sistema Digestivo.akospinaa100% (1)
- Practica 5. Epitelio GlandularDocumento17 páginasPractica 5. Epitelio GlandularNorma GL100% (1)
- Histología HépaticaDocumento4 páginasHistología HépaticaAngMag.Aún no hay calificaciones
- EFAp Exoc 10 Dr. García 2023-1Documento83 páginasEFAp Exoc 10 Dr. García 2023-1Mariana GamboaAún no hay calificaciones
- Epitelios GlandularesDocumento16 páginasEpitelios GlandularesAna MaríaAún no hay calificaciones
- Esofago y Tubo Digestivo-RpcDocumento7 páginasEsofago y Tubo Digestivo-RpcCarla RivasAún no hay calificaciones
- PancreasDocumento2 páginasPancreasd0ugy100% (3)
- CELULA GLANDULARrDocumento8 páginasCELULA GLANDULARrSantiago Andres Quishpe MartinezAún no hay calificaciones
- Glándulas SalivalesDocumento5 páginasGlándulas SalivalesAndrea Garcia100% (1)
- HIGADODocumento5 páginasHIGADOd0ugy100% (4)
- Fisiología de La Secreción SalivaDocumento23 páginasFisiología de La Secreción SalivaAidaLima100% (1)
- Actividad 11 Sistema Digestivo IIDocumento9 páginasActividad 11 Sistema Digestivo IIDaniela Estefania Garcia JuarezAún no hay calificaciones
- Usmp Practica 8 9 10 - Histologia 2013 - Aparato DigestivoDocumento119 páginasUsmp Practica 8 9 10 - Histologia 2013 - Aparato DigestivoCarito AlarconAún no hay calificaciones
- Bolillas ERA 3 HistoDocumento8 páginasBolillas ERA 3 HistoWolisarAún no hay calificaciones
- LO MAS IMPORTANTE CHyE 3Documento25 páginasLO MAS IMPORTANTE CHyE 3Ávila Simenoff MailénAún no hay calificaciones
- GLÁNDULASDocumento83 páginasGLÁNDULASDannaDoriaAún no hay calificaciones
- Esofago EstomagoDocumento4 páginasEsofago Estomagod0ugy100% (2)
- Apuntes Eptelios GlandularesDocumento13 páginasApuntes Eptelios GlandularesvalentinaAún no hay calificaciones
- Clase 6, GlandulasDocumento5 páginasClase 6, GlandulasLiz Mariel LizAún no hay calificaciones
- PAAF de HigadoDocumento7 páginasPAAF de HigadoRaquelAún no hay calificaciones
- HistologiaDocumento7 páginasHistologiaBenjamin AlvarezAún no hay calificaciones
- Rsumen para La InfografiaDocumento2 páginasRsumen para La InfografiaAlberto NavarroAún no hay calificaciones
- Glándulas SalivalesDocumento60 páginasGlándulas Salivalesalex79gfcgfAún no hay calificaciones
- GUIA de Epitelio GlandularDocumento22 páginasGUIA de Epitelio GlandularJessica0% (1)
- Las Glándulas SalivalesDocumento6 páginasLas Glándulas SalivalesLE LEAún no hay calificaciones
- Intoxicación Criminal Por GlifosatoDocumento25 páginasIntoxicación Criminal Por Glifosatomanuel herradoraAún no hay calificaciones
- Periodos Clinicos Del Parto.2023.Documento50 páginasPeriodos Clinicos Del Parto.2023.manuel herradoraAún no hay calificaciones
- Piel y HeridasDocumento51 páginasPiel y Heridasmanuel herradoraAún no hay calificaciones
- Presentación Grupal de Embarazo EctópicoDocumento12 páginasPresentación Grupal de Embarazo Ectópicomanuel herradoraAún no hay calificaciones
- Terminología QuirurgicaDocumento30 páginasTerminología Quirurgicamanuel herradoraAún no hay calificaciones
- Induccion ConduccionDocumento20 páginasInduccion Conduccionmanuel herradoraAún no hay calificaciones
- Criminalistica ForenseDocumento75 páginasCriminalistica Forensemanuel herradoraAún no hay calificaciones
- Its Electiva FinalDocumento95 páginasIts Electiva Finalmanuel herradoraAún no hay calificaciones
- EPOCDocumento5 páginasEPOCmanuel herradoraAún no hay calificaciones
- Antidepre 23Documento45 páginasAntidepre 23manuel herradoraAún no hay calificaciones
- Ecv DR Guzman ActualizadaDocumento108 páginasEcv DR Guzman Actualizadamanuel herradoraAún no hay calificaciones
- 7 Canal EndemicoDocumento25 páginas7 Canal Endemicomanuel herradoraAún no hay calificaciones
- Electiva Apn 2022Documento14 páginasElectiva Apn 2022manuel herradoraAún no hay calificaciones
- Anticonceptivos ElectivaDocumento91 páginasAnticonceptivos Electivamanuel herradoraAún no hay calificaciones
- Portafolio de Examen ObstetricoDocumento6 páginasPortafolio de Examen Obstetricomanuel herradoraAún no hay calificaciones
- Diabetes 23Documento24 páginasDiabetes 23manuel herradoraAún no hay calificaciones
- El Peritaje Forense AlacDocumento27 páginasEl Peritaje Forense Alacmanuel herradoraAún no hay calificaciones
- Cuadro Sinóptico de Llaves Simple Divertido y ColoridoDocumento1 páginaCuadro Sinóptico de Llaves Simple Divertido y Coloridomanuel herradoraAún no hay calificaciones
- Conferencia DigestiónDocumento33 páginasConferencia Digestiónmanuel herradoraAún no hay calificaciones
- Seminario Bioquimica 1Documento14 páginasSeminario Bioquimica 1manuel herradora0% (1)
- MALARIADocumento13 páginasMALARIAmanuel herradoraAún no hay calificaciones
- Ciencias Naturales (JoseDocumento22 páginasCiencias Naturales (Josemanuel herradoraAún no hay calificaciones
- Guia Practica Riesgo PsicDocumento3 páginasGuia Practica Riesgo Psicmanuel herradoraAún no hay calificaciones
- 4.1 - Diseños de Estudios EpidemilógicosDocumento46 páginas4.1 - Diseños de Estudios Epidemilógicosmanuel herradoraAún no hay calificaciones
- Balance HidromineralDocumento23 páginasBalance Hidromineralmanuel herradoraAún no hay calificaciones
- Pruebas TamizajeDocumento24 páginasPruebas Tamizajemanuel herradoraAún no hay calificaciones
- Diferenciacion 20 AnaplasiaDocumento2 páginasDiferenciacion 20 Anaplasiamanuel herradoraAún no hay calificaciones
- ATROFIADocumento3 páginasATROFIAmanuel herradoraAún no hay calificaciones
- Accidente de TráficoDocumento2 páginasAccidente de Tráficomanuel herradoraAún no hay calificaciones
- Microbiologia 1Documento11 páginasMicrobiologia 1manuel herradoraAún no hay calificaciones
- Resumen CazadorDocumento3 páginasResumen CazadorJuan Jesús Espinoza AmayaAún no hay calificaciones
- Amancaes GerminacionDocumento7 páginasAmancaes GerminacionAvia Solano SofíaAún no hay calificaciones
- 10-12 Problemas GenéticaDocumento3 páginas10-12 Problemas GenéticaAndrés Felipe Sánchez FernándezAún no hay calificaciones
- MODULO 1 (2) Pronap 2012 PDFDocumento86 páginasMODULO 1 (2) Pronap 2012 PDFNadia Savoye100% (1)
- La CélulaDocumento5 páginasLa Célular3750696Aún no hay calificaciones
- Asepsia y AntisepsiaDocumento24 páginasAsepsia y AntisepsiaRosangel RiveraAún no hay calificaciones
- Paradoja Del Plancton - ResumenDocumento1 páginaParadoja Del Plancton - Resumengilary peñaAún no hay calificaciones
- Carnes PavoDocumento6 páginasCarnes PavoAlbert Solis DiazAún no hay calificaciones
- 2 Tejido Epitelial 2010Documento50 páginas2 Tejido Epitelial 2010Jorge AguilarAún no hay calificaciones
- Formato de Artiuclos Facultad de Salud PublicaDocumento21 páginasFormato de Artiuclos Facultad de Salud PublicaGabyGarciaAún no hay calificaciones
- Manual Curso Buen Trato Laboral y Calidad de VidaDocumento46 páginasManual Curso Buen Trato Laboral y Calidad de VidaCristian Cortés RodríguezAún no hay calificaciones
- Ce RebeloDocumento39 páginasCe RebeloHugo DragoAún no hay calificaciones
- Trabajo FinalDocumento11 páginasTrabajo FinalyampierAún no hay calificaciones
- EnfermedadesInfecciosas Cto MexicoDocumento144 páginasEnfermedadesInfecciosas Cto MexicoAnonymous KZnCo5hv100% (2)
- Estructura y Elaboración de Microciclos de EntrenamientoDocumento27 páginasEstructura y Elaboración de Microciclos de Entrenamientofelisamt6996100% (5)
- Fundamento de Acuicultura Continental 3Documento266 páginasFundamento de Acuicultura Continental 3Diego Martínez CastroAún no hay calificaciones
- Tema 64Documento8 páginasTema 64Virginia Romero CarrascoAún no hay calificaciones
- Examen 2 ScientiaDocumento4 páginasExamen 2 ScientiaLeonardo Felipe Diestra Quiroz100% (1)
- Reservas Naturales de ArequipaDocumento2 páginasReservas Naturales de ArequipaPaolo AlessandroAún no hay calificaciones
- Ficha Tecnica In-ForlexDocumento2 páginasFicha Tecnica In-ForlexradacoAún no hay calificaciones
- Morfologia AlgodonDocumento4 páginasMorfologia AlgodonCristian WernerAún no hay calificaciones
- Proceso de Atun EnlatadoDocumento21 páginasProceso de Atun EnlatadoIsra Fragoso100% (1)
- Vademecum EdwardDocumento1 páginaVademecum Edwardedward paezAún no hay calificaciones
- Hemorragia en GestantesDocumento165 páginasHemorragia en GestantesIrving DTAún no hay calificaciones
- Define Tu Propósito de Vida en 3 HorasDocumento29 páginasDefine Tu Propósito de Vida en 3 HorasMario ChoqueticllaAún no hay calificaciones
- Resultadosindividualesi - 704505035 2Documento2 páginasResultadosindividualesi - 704505035 2Jennifer GamasAún no hay calificaciones
- Factores Etiológicos de Las CariesDocumento2 páginasFactores Etiológicos de Las CariesPablo Andrés Erazo100% (1)
- File 1599515788Documento13 páginasFile 1599515788juan pablo escandón leguizamoAún no hay calificaciones
- Agentes LeudantesDocumento9 páginasAgentes LeudantesPaul Andres Segovia Macias100% (8)
- Sistema Digestivo y RenalDocumento5 páginasSistema Digestivo y RenalAleAún no hay calificaciones