También podría gustarte
- Sistema de complemento: funciones y activaciónDocumento11 páginasSistema de complemento: funciones y activaciónIngrid Tatiana LozanoAún no hay calificaciones
- Sistema complementoDocumento5 páginasSistema complementoLolaAún no hay calificaciones
- Sistema Del ComplementoDocumento8 páginasSistema Del ComplementoPaula VelezAún no hay calificaciones
- Sistemas de Complemento: Cascadas BioquímicasDocumento3 páginasSistemas de Complemento: Cascadas BioquímicasCamzAún no hay calificaciones
- Repaso tema 4 inmunologiaDocumento8 páginasRepaso tema 4 inmunologiaorlynmagdalysAún no hay calificaciones
- INMUNOLOGIA APUNTES3pDocumento8 páginasINMUNOLOGIA APUNTES3pLuis FernandoAún no hay calificaciones
- Guía Práctica de Complemento Septiembre 2011Documento7 páginasGuía Práctica de Complemento Septiembre 2011InmunoBlog100% (1)
- El Sistema Del ComplementoDocumento7 páginasEl Sistema Del ComplementojesusAún no hay calificaciones
- Sistema Del ComplementoDocumento14 páginasSistema Del ComplementoDeysi LévanoAún no hay calificaciones
- El Sistema Del ComplementoDocumento5 páginasEl Sistema Del ComplementoPAOLA ARCILAAún no hay calificaciones
- Teoria 6 InmunoDocumento16 páginasTeoria 6 Inmunoandrea lyaAún no hay calificaciones
- Resumen Via AlternaDocumento8 páginasResumen Via AlternaPRISGPEAún no hay calificaciones
- Sistema del complemento: ligandos, receptores y vías de activaciónDocumento5 páginasSistema del complemento: ligandos, receptores y vías de activaciónIreneSanzGilAún no hay calificaciones
- Sistema Del ComplementoDocumento6 páginasSistema Del ComplementoDiego JimenezAún no hay calificaciones
- Sistema del complementoDocumento6 páginasSistema del complementomicaelassciAún no hay calificaciones
- .Sistema de Complemento .Documento11 páginas.Sistema de Complemento .Janacua Figueroa DianaAún no hay calificaciones
- EL Sistema DEL ComplementoDocumento5 páginasEL Sistema DEL ComplementoMiranda LeonettiAún no hay calificaciones
- Sistema de Complemento - IfDocumento9 páginasSistema de Complemento - IfManuel Alejandro Lengua CuliAún no hay calificaciones
- Componentes Del ComplementoDocumento4 páginasComponentes Del ComplementonaddhxielyAún no hay calificaciones
- ComplementoDocumento9 páginasComplementopukka38Aún no hay calificaciones
- 5 Sistema de ComplementoDocumento40 páginas5 Sistema de ComplementoLuan NettoAún no hay calificaciones
- + Sistema Del ComplementoDocumento6 páginas+ Sistema Del ComplementoAntonella DuarteAún no hay calificaciones
- Sistema de ComlementoDocumento24 páginasSistema de ComlementoEdwar Leyva BaltazarAún no hay calificaciones
- Sistema de ComplementoDocumento17 páginasSistema de ComplementoCarlosErazoAún no hay calificaciones
- Sistema Del Complemento - Inmunología y Trastornos Alérgicos - Manual MSD Versión para ProfesionalesDocumento2 páginasSistema Del Complemento - Inmunología y Trastornos Alérgicos - Manual MSD Versión para ProfesionalesWilliam Aarón Vargas SalmerónAún no hay calificaciones
- 194 11440 1 PBDocumento9 páginas194 11440 1 PB2mf2hgjrj9Aún no hay calificaciones
- El Sistema Del ComplementoDocumento3 páginasEl Sistema Del ComplementoYuliana PereiraAún no hay calificaciones
- Sistema Del ComplementoDocumento12 páginasSistema Del ComplementoSewa FloresAún no hay calificaciones
- SistemadelComplemento CREDocumento5 páginasSistemadelComplemento CRERec 379Aún no hay calificaciones
- ComplementoDocumento10 páginasComplementoGomez UrsuAún no hay calificaciones
- Sistema del complemento: vías de activación, receptores y funcionesDocumento45 páginasSistema del complemento: vías de activación, receptores y funcionesYasmin FigueroaAún no hay calificaciones
- Sistema de Complemento (Resumen)Documento16 páginasSistema de Complemento (Resumen)Edgar D Sanchez OliverosAún no hay calificaciones
- Cuestionario Artículo de S. ComplementoDocumento3 páginasCuestionario Artículo de S. ComplementoVeronicaRubíJAún no hay calificaciones
- Sistema de Complemento: Definición, Características y VíasDocumento12 páginasSistema de Complemento: Definición, Características y VíasMaría Alvarado PachasAún no hay calificaciones
- Sistema de ComplementoDocumento27 páginasSistema de ComplementoBiali LopezAún no hay calificaciones
- Sistema de Complemento (RESUMEN)Documento16 páginasSistema de Complemento (RESUMEN)Edgar D Sanchez OliverosAún no hay calificaciones
- Clase 4-23 TMDocumento73 páginasClase 4-23 TMYuLi WAún no hay calificaciones
- Cuestionario Del Sistema Del ComplementoDocumento6 páginasCuestionario Del Sistema Del ComplementoKarla BastidaAún no hay calificaciones
- Actualización sobre el complemento y sus vías de activaciónDocumento12 páginasActualización sobre el complemento y sus vías de activacióntere ariasAún no hay calificaciones
- Inmunidad y AntibioticosDocumento13 páginasInmunidad y AntibioticosKrystel Volgger GarciaAún no hay calificaciones
- Ensayo Del Sistema de ComplementoDocumento2 páginasEnsayo Del Sistema de ComplementoKevin Denis ChoqueAún no hay calificaciones
- Sistema de ComplementoDocumento20 páginasSistema de ComplementoAngel CastilloAún no hay calificaciones
- FagocitocisDocumento4 páginasFagocitocisRai GuerreroAún no hay calificaciones
- Complemento Mapa ConceptualDocumento14 páginasComplemento Mapa ConceptualJDominguezBernalAún no hay calificaciones
- Sistema Del ComplementoDocumento35 páginasSistema Del ComplementoRomina ArgumedoAún no hay calificaciones
- Lecturas 2do Parcial Regueiro InmunologíaDocumento16 páginasLecturas 2do Parcial Regueiro Inmunologíaanaca59505Aún no hay calificaciones
- Sistema de ComplementoDocumento13 páginasSistema de ComplementoKaren Estefany Rengifo CruzadoAún no hay calificaciones
- Sist. ComplementoDocumento9 páginasSist. ComplementoELENA YAPURA BILBAOAún no hay calificaciones
- 6-Sistema de ComplementoDocumento9 páginas6-Sistema de Complementogeobannys cedeñoAún no hay calificaciones
- Carpeta Semana 6Documento4 páginasCarpeta Semana 6ISELLE SUERO MATOSAún no hay calificaciones
- Sistema de Complemento, Vias y FuncionesDocumento6 páginasSistema de Complemento, Vias y FuncionesAdrian Briones100% (1)
- El Sistema Del ComplementoDocumento5 páginasEl Sistema Del Complementovictor quispe CastroAún no hay calificaciones
- Complemento y CininasDocumento8 páginasComplemento y CininasCarlos RochaAún no hay calificaciones
- Sistema de ComplementoDocumento9 páginasSistema de ComplementoLauraCarolinaRestrepoAún no hay calificaciones
- Sistema de ComplementoDocumento8 páginasSistema de Complementonathaly barretoAún no hay calificaciones
- Sistema de Complemento ModificadoDocumento14 páginasSistema de Complemento ModificadoYoshiro ReañoAún no hay calificaciones
- Inflamacion, Fagocitosis y ComplementoDocumento49 páginasInflamacion, Fagocitosis y ComplementoNayeli Espinosa0% (1)
- Sistema de Complemento: Funciones y NomenclaturaDocumento11 páginasSistema de Complemento: Funciones y NomenclaturaAna Jose Viveros HigueraAún no hay calificaciones
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- Práctica 1. Equipo 10Documento9 páginasPráctica 1. Equipo 10Ale SaldivarAún no hay calificaciones
- Práctica 3. Equipo 10Documento12 páginasPráctica 3. Equipo 10Ale SaldivarAún no hay calificaciones
- Práctica 2. Equipo 10Documento10 páginasPráctica 2. Equipo 10Ale SaldivarAún no hay calificaciones
- Prácitica 5. Equipo 10Documento15 páginasPrácitica 5. Equipo 10Ale SaldivarAún no hay calificaciones
- Práctica 0. Equipo 10Documento12 páginasPráctica 0. Equipo 10Ale SaldivarAún no hay calificaciones
- InmunodeficienciasDocumento12 páginasInmunodeficienciasAle SaldivarAún no hay calificaciones
- LignanosDocumento5 páginasLignanosAle SaldivarAún no hay calificaciones
- Práctica 4. Equipo 10Documento10 páginasPráctica 4. Equipo 10Ale SaldivarAún no hay calificaciones
- Perspectiva de Los FitomedicinalesDocumento4 páginasPerspectiva de Los FitomedicinalesAle SaldivarAún no hay calificaciones
- Sistema Músculo-EsqueléticoDocumento6 páginasSistema Músculo-EsqueléticoAle Saldivar100% (1)
- Observación de Las Células Del SIDocumento12 páginasObservación de Las Células Del SIAle SaldivarAún no hay calificaciones
- Estructura y función de los cromosomas en el ciclo celularDocumento22 páginasEstructura y función de los cromosomas en el ciclo celularAle SaldivarAún no hay calificaciones
- De Las MutacionesDocumento10 páginasDe Las MutacionesAle SaldivarAún no hay calificaciones
- De La Genética y El ADNDocumento15 páginasDe La Genética y El ADNAle SaldivarAún no hay calificaciones
- De La Ley de MendelDocumento9 páginasDe La Ley de MendelAle SaldivarAún no hay calificaciones
- De La Herencia No Clásica y MendelianaDocumento9 páginasDe La Herencia No Clásica y MendelianaAle SaldivarAún no hay calificaciones
- Niveles de OrganizaciónDocumento2 páginasNiveles de OrganizaciónAle SaldivarAún no hay calificaciones
- Reporte 04Documento9 páginasReporte 04Ale SaldivarAún no hay calificaciones
- Generalidades de Anatomía y FisiologíaDocumento2 páginasGeneralidades de Anatomía y FisiologíaAle SaldivarAún no hay calificaciones
- Obtención de N-OctanolDocumento5 páginasObtención de N-OctanolAle SaldivarAún no hay calificaciones
- Problemario 4 - DNA y RNADocumento2 páginasProblemario 4 - DNA y RNAAle SaldivarAún no hay calificaciones
- Problemario 01Documento3 páginasProblemario 01Ale SaldivarAún no hay calificaciones
- Reporte 06Documento6 páginasReporte 06Ale SaldivarAún no hay calificaciones
- Problemario Análisis InstrumentalDocumento12 páginasProblemario Análisis InstrumentalAle Saldivar100% (1)
- Proyecto de AstronomíaDocumento30 páginasProyecto de AstronomíaAle SaldivarAún no hay calificaciones
- Ibu MilenaDocumento18 páginasIbu MilenaGelfioAún no hay calificaciones
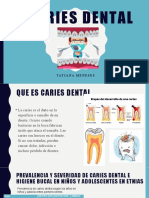
- Caries dental en niños y adolescentesDocumento5 páginasCaries dental en niños y adolescentesTatiana MenesesAún no hay calificaciones
- Repertorio de Habilidades Pre-AcademicasDocumento6 páginasRepertorio de Habilidades Pre-AcademicasDulce Fuentes0% (1)
- Informe Técnico Área Psicología Clínica Del Hospital Especializado San Juan de DiosDocumento4 páginasInforme Técnico Área Psicología Clínica Del Hospital Especializado San Juan de DiosAndrés ParraAún no hay calificaciones
- Folleto Riesgo QuimicoDocumento3 páginasFolleto Riesgo QuimicoLuisAún no hay calificaciones
- TRIPTICODocumento3 páginasTRIPTICOPachecomx PachecoAún no hay calificaciones
- TraqueostomíaDocumento17 páginasTraqueostomíaFranco MamaniAún no hay calificaciones
- Fundamentos de la clínica psicológica y el inconsciente según FreudDocumento46 páginasFundamentos de la clínica psicológica y el inconsciente según FreudElle BlackAún no hay calificaciones
- Las Hormonas Que Intervienen en El Ciclo MenstrualDocumento2 páginasLas Hormonas Que Intervienen en El Ciclo MenstrualMaría José Vásquez TarrilloAún no hay calificaciones
- RCO - N - 112-2022-UNTELS - Directiva para El Retorno Al Trabajo en La Modalidad Presencial o Mixta UNTELSDocumento12 páginasRCO - N - 112-2022-UNTELS - Directiva para El Retorno Al Trabajo en La Modalidad Presencial o Mixta UNTELSMirtha Ochoa GamarraAún no hay calificaciones
- Intervencion Breve para CocaínaDocumento62 páginasIntervencion Breve para CocaínaAdriana Ornelas AguirreAún no hay calificaciones
- Actividad física escuela saludDocumento23 páginasActividad física escuela saludmariaAún no hay calificaciones
- La Alexia - DislexiaDocumento23 páginasLa Alexia - DislexiaPedro Luis Salas AchateAún no hay calificaciones
- Hyaluron Pen - Nano PenDocumento70 páginasHyaluron Pen - Nano PenLya Silva100% (1)
- Info 10 Es023 Epidemiología SemDocumento6 páginasInfo 10 Es023 Epidemiología SemAleysi Cieza DelgadoAún no hay calificaciones
- ELISA Es Un Mens@ (Final, Version Chistosa Sin Video)Documento35 páginasELISA Es Un Mens@ (Final, Version Chistosa Sin Video)Karla Perez CuhuatecatlAún no hay calificaciones
- Evaluación final SST introducciónDocumento7 páginasEvaluación final SST introducciónAngela CaballeroAún no hay calificaciones
- Notificaciones de Accidentes de Trabajo, Incidentes Peligrosos y Enfermedades OcupacionalesDocumento29 páginasNotificaciones de Accidentes de Trabajo, Incidentes Peligrosos y Enfermedades OcupacionalesRudy Enryque Quiroz100% (1)
- Clinica Pancreatitis AgudaDocumento28 páginasClinica Pancreatitis AgudaSantiago JoseAún no hay calificaciones
- El niño en contexto social y familiarDocumento87 páginasEl niño en contexto social y familiarDulce HaquetAún no hay calificaciones
- Higienes y Seguridad IndustrialDocumento8 páginasHigienes y Seguridad IndustrialSANDRA NATALIA ACOSTA PARDOAún no hay calificaciones
- LuuuuzDocumento6 páginasLuuuuzYaricza Dayanara Orozco FloresAún no hay calificaciones
- E EXTREMO 2018 - Ginecología Obstetricia - Online PDFDocumento71 páginasE EXTREMO 2018 - Ginecología Obstetricia - Online PDFLichi Flores PalaciosAún no hay calificaciones
- Sordera o Perdida Auditiva-Lottys Aguilar.Documento1 páginaSordera o Perdida Auditiva-Lottys Aguilar.Lottys AguilarAún no hay calificaciones
- Total Result 1448: Edad - All - Genero Cuenta de #Cedula Femenino 918 Masculino 530Documento651 páginasTotal Result 1448: Edad - All - Genero Cuenta de #Cedula Femenino 918 Masculino 530blue starsAún no hay calificaciones
- Sepsis y Choque Septico.Documento20 páginasSepsis y Choque Septico.Jessica Cecilia Buenfil ArceoAún no hay calificaciones
- Efectos de La Moxa en El Recuento Plaquetario HumanoDocumento45 páginasEfectos de La Moxa en El Recuento Plaquetario HumanoLINOAún no hay calificaciones
- Historia ClínicaDocumento3 páginasHistoria ClínicaRicardo AndradeAún no hay calificaciones
- RUC .20154489895 Dirección: Calle El Filtro 501 Distrito: Cercado Provincia: Arequipa Departamento: Arequipa Ii. Datos de Lugares de TrabajoDocumento33 páginasRUC .20154489895 Dirección: Calle El Filtro 501 Distrito: Cercado Provincia: Arequipa Departamento: Arequipa Ii. Datos de Lugares de TrabajoErika LionetAún no hay calificaciones
- Guía Práctica LG 2022Documento5 páginasGuía Práctica LG 2022Larisa VillanuevaAún no hay calificaciones