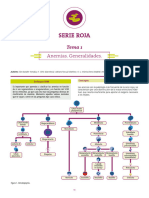
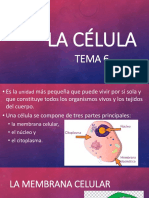
También podría gustarte
- Informe 3 GravimetríaDocumento6 páginasInforme 3 GravimetríaLaura SánchezAún no hay calificaciones
- FenazopiridinaDocumento7 páginasFenazopiridinaAnonymous ToMMmIv100% (1)
- Informe de Soluciones BufferDocumento11 páginasInforme de Soluciones BufferRoy Brañez0% (3)
- Intoxicación Por SalicilatosDocumento4 páginasIntoxicación Por Salicilatoscarmen celenia barrios quirozAún no hay calificaciones
- Caso Clínico N2. Insuficiencia CardíacaDocumento44 páginasCaso Clínico N2. Insuficiencia CardíacamiltonAún no hay calificaciones
- Ferrum16 - Multivitaminico para Toda La Familia...Documento22 páginasFerrum16 - Multivitaminico para Toda La Familia...Ancalmo InternacionalAún no hay calificaciones
- Practica # 6 - Fosfatasa AlcalinaDocumento7 páginasPractica # 6 - Fosfatasa Alcalinarockakicha100% (1)
- Determinacion Cuantitativa Del Acido UricoDocumento14 páginasDeterminacion Cuantitativa Del Acido UricoPedro SuarezAún no hay calificaciones
- Semana 10 - Toxicologia y Quimica Legal - Guia de Practica - Det Barbituricos - Benzodiazepias y FenotiazinicosDocumento8 páginasSemana 10 - Toxicologia y Quimica Legal - Guia de Practica - Det Barbituricos - Benzodiazepias y FenotiazinicosLinsay GranadosAún no hay calificaciones
- PREGUNTASDocumento5 páginasPREGUNTASyeniAún no hay calificaciones
- Farmacos adrenergicos y colinergicos en frecuencia cardiaca y respiratoriaDocumento15 páginasFarmacos adrenergicos y colinergicos en frecuencia cardiaca y respiratoriaWilma Valiente TantaAún no hay calificaciones
- Determinación de Compuestos NitrogenadosDocumento8 páginasDeterminación de Compuestos NitrogenadosLourdes TapuyAún no hay calificaciones
- Listado de Medicamentos de Estrecho Margen Terapeuticos PDFDocumento5 páginasListado de Medicamentos de Estrecho Margen Terapeuticos PDFJorge Ivan Patiño CuervoAún no hay calificaciones
- Aguilar Arteaga 8Documento5 páginasAguilar Arteaga 8VANESSA ALEXANDRA AGUILAR ARTEAGAAún no hay calificaciones
- 2) Neuroepilepticos AntimaniacosDocumento10 páginas2) Neuroepilepticos AntimaniacosLucero Montes HuanquiAún no hay calificaciones
- PRÁCTICA No - 1Documento3 páginasPRÁCTICA No - 1manuel torresAún no hay calificaciones
- Aspirina InformeDocumento20 páginasAspirina InformeEduardo Poma100% (1)
- Farmacoquímica examen finalDocumento4 páginasFarmacoquímica examen finalWindows 7Aún no hay calificaciones
- Cuestionario Bioquimica Clinica WraDocumento10 páginasCuestionario Bioquimica Clinica WraWara OrellanaAún no hay calificaciones
- Ejercicios QuimicaDocumento7 páginasEjercicios QuimicaMelisa RodriguezAún no hay calificaciones
- Cuestionario de La Practica de LipidosDocumento5 páginasCuestionario de La Practica de LipidosErik Ortiz BlancoAún no hay calificaciones
- Gases ideales: tipos y propiedadesDocumento2 páginasGases ideales: tipos y propiedadesAdriana Jasso RuizAún no hay calificaciones
- Factores de riesgo Sildenafil adolescentesDocumento43 páginasFactores de riesgo Sildenafil adolescentesyolaymi peralta100% (1)
- Taller 1 Bibliografia de Cientificos y Su Aporte en Ana. InstrumentalDocumento4 páginasTaller 1 Bibliografia de Cientificos y Su Aporte en Ana. InstrumentalPaul Vargas100% (1)
- Análisis bromatológico de agua de consumoDocumento9 páginasAnálisis bromatológico de agua de consumoKaterin Rondo TolentinoAún no hay calificaciones
- Glicerolado para grietasDocumento6 páginasGlicerolado para grietasLuiz MiguelAún no hay calificaciones
- Informe CoDocumento8 páginasInforme CoYsamar Tapia BañezAún no hay calificaciones
- Laboratorio BilirubinaDocumento17 páginasLaboratorio BilirubinaAlejandra BohórquezAún no hay calificaciones
- Glucosa Hexoquinasa PDFDocumento2 páginasGlucosa Hexoquinasa PDFFujiko Saavedra Leiva100% (2)
- ANON-Apuntes y Lecciones de Química Farmaceútica, Universidad de AlcaláDocumento133 páginasANON-Apuntes y Lecciones de Química Farmaceútica, Universidad de AlcaláJuanManuelAmaroLuisAún no hay calificaciones
- Desproteinizados y Ácido ÚricoDocumento36 páginasDesproteinizados y Ácido ÚricoMarcelo MendezAún no hay calificaciones
- Determinación de salicilatos en orinaDocumento10 páginasDeterminación de salicilatos en orinaAriana MuñozAún no hay calificaciones
- Ficha Tecnica Sodio Almidon Glicolato FagronDocumento1 páginaFicha Tecnica Sodio Almidon Glicolato FagronportosinAún no hay calificaciones
- Practica 15 MarihuanaDocumento8 páginasPractica 15 MarihuanaBilly Joe Narvaez RamosAún no hay calificaciones
- Práctica 10 Ácido Úrico y GotaDocumento8 páginasPráctica 10 Ácido Úrico y GotaVICTORIA KATHERINE RODRIGUEZ VALLE100% (1)
- Informe de SalbutamolDocumento14 páginasInforme de SalbutamolWiny Yucra Chaqquere100% (2)
- Extracción de lactonas sesquiterpénicas en la alcachofaDocumento3 páginasExtracción de lactonas sesquiterpénicas en la alcachofaCristhian P. RojasAún no hay calificaciones
- Prcticas de Lab. Organica IiDocumento20 páginasPrcticas de Lab. Organica IiGabriel Guzman Martinez100% (1)
- Control de Síntomas en Paciente Terminal - Diana Zelada MallcoDocumento25 páginasControl de Síntomas en Paciente Terminal - Diana Zelada MallcoEliana BravoAún no hay calificaciones
- Determinanación Por Espectrofotometría El Contenido de La Glucosa en Suero Sanguíneo HumanoDocumento4 páginasDeterminanación Por Espectrofotometría El Contenido de La Glucosa en Suero Sanguíneo HumanogravisAún no hay calificaciones
- Identificacion de Antocianidinas PRACTICA 11Documento8 páginasIdentificacion de Antocianidinas PRACTICA 11Zara ClarosAún no hay calificaciones
- Informe-Analisis Bromatologico de AguasDocumento13 páginasInforme-Analisis Bromatologico de AguasDennys Yamer Cueva TandaipanAún no hay calificaciones
- Metabolismo de Acido UricoDocumento2 páginasMetabolismo de Acido UricoAntonio PiñaAún no hay calificaciones
- 27 Lipidos Simples 2019Documento52 páginas27 Lipidos Simples 2019Fernanda Gutiérrez EscobarAún no hay calificaciones
- Bolus IV. Modelo Monocompartimental AbiertoDocumento15 páginasBolus IV. Modelo Monocompartimental AbiertoLuis Alberto LRAún no hay calificaciones
- Anticoagulantes in VivoDocumento39 páginasAnticoagulantes in VivobAún no hay calificaciones
- Laboratorio N°7 - ProteínasDocumento4 páginasLaboratorio N°7 - ProteínaseleazarAún no hay calificaciones
- Intoxicación por plaguicidas y tratamiento de íleo paralíticoDocumento3 páginasIntoxicación por plaguicidas y tratamiento de íleo paralíticoEduardo Alejandro Cedeno EspinozaAún no hay calificaciones
- Caso Clinico 1 OficialDocumento52 páginasCaso Clinico 1 OficialDiana Diaz100% (1)
- Práctica N°7 - Fármacos Estimulantes Del Sistema Nervioso CentralDocumento5 páginasPráctica N°7 - Fármacos Estimulantes Del Sistema Nervioso CentralRUTH GABRIELA BENAVIDEZ CARBAJALAún no hay calificaciones
- Analisis Bromatologico de VinosDocumento9 páginasAnalisis Bromatologico de VinosKaterin Rondo TolentinoAún no hay calificaciones
- Agua BoricadaDocumento4 páginasAgua Boricadacarlosmax5_55Aún no hay calificaciones
- Materia OrgánicaDocumento33 páginasMateria OrgánicaDanny Gabriel Diaz NajarAún no hay calificaciones
- AntibióticosDocumento9 páginasAntibióticosAdriana MXAún no hay calificaciones
- PLANILLA Subestacion ElectricaDocumento10 páginasPLANILLA Subestacion Electricaruben colqueAún no hay calificaciones
- Proced I Mien ToDocumento3 páginasProced I Mien ToMayraSamameBurgosAún no hay calificaciones
- Acidos FosfatidicosDocumento8 páginasAcidos FosfatidicosMatthew BrooksAún no hay calificaciones
- Fármacos Que Actúan Sobre Receptores Intracelulares WordDocumento13 páginasFármacos Que Actúan Sobre Receptores Intracelulares WordD Lorena Torres100% (2)
- Cuestionario 12Documento4 páginasCuestionario 12usmpusmpusmp100% (1)
- Ramirez - Investigación LipidosDocumento6 páginasRamirez - Investigación LipidosRAMIRO JOSE RAMIREZ ROMEROAún no hay calificaciones
- Clase 4 Medicina Colonial UltimoDocumento59 páginasClase 4 Medicina Colonial UltimoSamuel FloresAún no hay calificaciones
- GRECIADocumento55 páginasGRECIAcarlos miguelAún no hay calificaciones
- CLASE 5 Finales Colonia Hipolito UnanueDocumento45 páginasCLASE 5 Finales Colonia Hipolito Unanuecarlos miguelAún no hay calificaciones
- ASTROLOGÍADocumento26 páginasASTROLOGÍAcarlos miguelAún no hay calificaciones
- Peru PrehispanicoDocumento44 páginasPeru Prehispanicocarlos miguelAún no hay calificaciones
- CLASE 5 Finales Colonia Hipolito UnanueDocumento45 páginasCLASE 5 Finales Colonia Hipolito Unanuecarlos miguelAún no hay calificaciones
- Clase 1 Historia de La MedicinaDocumento32 páginasClase 1 Historia de La MedicinaLuigiZeladaNeyraAún no hay calificaciones
- Origen Del Hombre UltimoDocumento49 páginasOrigen Del Hombre Ultimocarlos miguelAún no hay calificaciones
- Clase 1 Historia de La MedicinaDocumento32 páginasClase 1 Historia de La MedicinaLuigiZeladaNeyraAún no hay calificaciones
- Informe Practica Semana 11Documento12 páginasInforme Practica Semana 11carlos miguelAún no hay calificaciones
- Diabetes - TratamientoDocumento6 páginasDiabetes - Tratamientocarlos miguelAún no hay calificaciones
- Informe de EmbriologiaDocumento29 páginasInforme de Embriologiacarlos miguelAún no hay calificaciones
- Informe de Embriologia FinalDocumento59 páginasInforme de Embriologia Finalcarlos miguelAún no hay calificaciones
- Informe Bases MolecularesDocumento19 páginasInforme Bases Molecularescarlos miguelAún no hay calificaciones
- Informe de Bases MolecularesDocumento13 páginasInforme de Bases Molecularescarlos miguelAún no hay calificaciones
- Tipos de Investigación Según Su PropositoDocumento5 páginasTipos de Investigación Según Su Propositocarlos miguel100% (1)
- Tipos de células: procariotas vs eucariotasDocumento5 páginasTipos de células: procariotas vs eucariotasSanchez Salluca Gian MarcoAún no hay calificaciones
- Factor de Seguridad para La Transmisión en La Unión NeuromuscularDocumento4 páginasFactor de Seguridad para La Transmisión en La Unión NeuromuscularObstetras Diego VictoriaAún no hay calificaciones
- Informe #3 Microbiología Sedimentación y Coloración de GramDocumento5 páginasInforme #3 Microbiología Sedimentación y Coloración de Gramyenny lorena trujillo trujilloAún no hay calificaciones
- Neurona TrabajoDocumento9 páginasNeurona TrabajoMalvina Soledad TorresAún no hay calificaciones
- CalicreínaDocumento12 páginasCalicreínaCinthya Merma CcanaAún no hay calificaciones
- AminoacidosDocumento3 páginasAminoacidosCarlos BrownAún no hay calificaciones
- Leucina zipper regula factores de transcripciónDocumento2 páginasLeucina zipper regula factores de transcripciónLOL X Repleys ElmoshtaykersAún no hay calificaciones
- Unidad 1Documento125 páginasUnidad 1JULIO CESAR GUTIERREZ CONDORIAún no hay calificaciones
- Hematologia: Laboratorio Lorena Vejarano S.ADocumento2 páginasHematologia: Laboratorio Lorena Vejarano S.ALau JaramilloAún no hay calificaciones
- Sistema Inmunológico 1 oryl3FSDocumento14 páginasSistema Inmunológico 1 oryl3FSOmayra MartínezAún no hay calificaciones
- Organelos Celulares y Sus FuncionesDocumento22 páginasOrganelos Celulares y Sus FuncionesAngee BojorquezAún no hay calificaciones
- Embriogénesis y fecundación: procesos clave de la reproducción humanaDocumento31 páginasEmbriogénesis y fecundación: procesos clave de la reproducción humanaMechePameAún no hay calificaciones
- Pp3 Mitosis y Meiosis 2medio AA - CCDocumento2 páginasPp3 Mitosis y Meiosis 2medio AA - CCJimena Gallo CaderónAún no hay calificaciones
- BIOLOGIA CLASE 4 CelulaDocumento1 páginaBIOLOGIA CLASE 4 CelulaPedro Vega QuispeAún no hay calificaciones
- Consolidación de Tegumentario CorregidoDocumento12 páginasConsolidación de Tegumentario CorregidoSangría MA&N2Aún no hay calificaciones
- Procesamiento Del ARNDocumento194 páginasProcesamiento Del ARNthiagoscathcartAún no hay calificaciones
- PielDocumento30 páginasPielClash SepAún no hay calificaciones
- IDs Primarias y Secundarias: causas, tipos y SIDADocumento3 páginasIDs Primarias y Secundarias: causas, tipos y SIDAMaria PerezAún no hay calificaciones
- 1579-Texto Del Artículo-5421-1-10-20220615Documento11 páginas1579-Texto Del Artículo-5421-1-10-20220615darbaderAún no hay calificaciones
- Hematología AMIR 14va Edición-13-36Documento24 páginasHematología AMIR 14va Edición-13-36Mauro VelascoAún no hay calificaciones
- La Célula-Teoria - Tipos de ReproducciónDocumento22 páginasLa Célula-Teoria - Tipos de ReproducciónjesicaAún no hay calificaciones
- La FototransducciónDocumento9 páginasLa FototransducciónJerry Carhuatanta SaavedraAún no hay calificaciones
- Practica VI BiotecnologiaDocumento26 páginasPractica VI BiotecnologiaHector Joel Chaname Rodriguez100% (1)
- PRÁCTICA No.7 Membrana y BioenergéticaDocumento11 páginasPRÁCTICA No.7 Membrana y BioenergéticaGabriela LpezAún no hay calificaciones
- Uniones celulares en epitelio intestinalDocumento14 páginasUniones celulares en epitelio intestinalIdaLii SancheezAún no hay calificaciones
- Adminpujojs, 08 TRANSICION EPITELIODocumento25 páginasAdminpujojs, 08 TRANSICION EPITELIOgg2022073533Aún no hay calificaciones
- Exposicion de Eritrocito FinalDocumento35 páginasExposicion de Eritrocito FinalGemhma CastellanosAún no hay calificaciones
- ResumenDocumento118 páginasResumenKaren Ospina SalazarAún no hay calificaciones
- Cta4-u2-Sesion 02 - Sesión Práctica de Laboratorio Observacion CelulaDocumento3 páginasCta4-u2-Sesion 02 - Sesión Práctica de Laboratorio Observacion CelulaShirley Pinedo100% (3)
- Cinética enzimática: conceptos básicosDocumento25 páginasCinética enzimática: conceptos básicosTonyFontanaAún no hay calificaciones