También podría gustarte
- Informe Del Proyecto Feto de BovinoDocumento13 páginasInforme Del Proyecto Feto de BovinoberthajachoAún no hay calificaciones
- Citología ClaseDocumento3 páginasCitología ClasesalazarthisAún no hay calificaciones
- RESUMEN Del Sisatema Alimentario EmbriologiaDocumento3 páginasRESUMEN Del Sisatema Alimentario EmbriologiakerlyAún no hay calificaciones
- EFAp Digest UPSJB III CICLODocumento187 páginasEFAp Digest UPSJB III CICLOBecky Betsy Burga GutierrezAún no hay calificaciones
- Resumen - Desarrollo Embrionario Del Sistema Gástrico y Sistema PulmonarDocumento5 páginasResumen - Desarrollo Embrionario Del Sistema Gástrico y Sistema PulmonarMayra MarroquinAún no hay calificaciones
- 12° Aparato DigestivoDocumento8 páginas12° Aparato DigestivoCarlos RomeroAún no hay calificaciones
- Desarrollo Embrionario Glandulas AnexasDocumento6 páginasDesarrollo Embrionario Glandulas Anexasgonzalo CHINOAún no hay calificaciones
- Embriología Del Sistema DigestivoDocumento4 páginasEmbriología Del Sistema DigestivoMilagros NuñezAún no hay calificaciones
- Sistema DigestDocumento6 páginasSistema DigestSol Arteaga RoaAún no hay calificaciones
- Para La Teoria de Morfo El 25Documento3 páginasPara La Teoria de Morfo El 25Taty GueledelAún no hay calificaciones
- Desarrollo Intestino PrimitivoDocumento74 páginasDesarrollo Intestino PrimitivoSarahi MercadoAún no hay calificaciones
- Aparato DigestivoDocumento10 páginasAparato DigestivoKamila Castillo VegaAún no hay calificaciones
- Embriologia Sistema DigestivoDocumento8 páginasEmbriologia Sistema DigestivoKatia FelizzolaAún no hay calificaciones
- Sistema digestivo embriologíaDocumento6 páginasSistema digestivo embriologíasandra acostaAún no hay calificaciones
- Saco VitelinoDocumento3 páginasSaco VitelinoJACINTO ROSALES YULIZA ZARAIAún no hay calificaciones
- Aparato DigestivoDocumento4 páginasAparato DigestivoAlejandro RSAún no hay calificaciones
- Seminario 3 EmbriologíaDocumento32 páginasSeminario 3 EmbriologíaMarco Francesco Barboza AraujoAún no hay calificaciones
- Cuestionario 4 y 5 - Derivados MesodermalesDocumento6 páginasCuestionario 4 y 5 - Derivados MesodermalesLucy Ascona100% (1)
- Desarrollo del aparato digestivo y respiratorioDocumento10 páginasDesarrollo del aparato digestivo y respiratorioMajo MVAún no hay calificaciones
- Desarrollo Del Sistema GastrointestinalDocumento56 páginasDesarrollo Del Sistema GastrointestinalNeydi MendezAún no hay calificaciones
- Aparato DigestivoDocumento10 páginasAparato DigestivoJoyce ZambranoAún no hay calificaciones
- 10.sistema DigestivoDocumento35 páginas10.sistema DigestivoLuis Alberto Benitez100% (2)
- Aparato Digestivo 2 Embrio 2023Documento4 páginasAparato Digestivo 2 Embrio 2023RaoulAún no hay calificaciones
- Cuestionario 3 - III UnidadDocumento9 páginasCuestionario 3 - III UnidadKatherine Mirella Saldaña CamposAún no hay calificaciones
- Embriologia Sistema DigestivoDocumento3 páginasEmbriologia Sistema DigestivoLisandra BerroaAún no hay calificaciones
- Desarrollo Del Aparato DigestivoDocumento11 páginasDesarrollo Del Aparato Digestivorafael mendezAún no hay calificaciones
- Embriologia Aparato 150756 Downloadable 4573836Documento11 páginasEmbriologia Aparato 150756 Downloadable 4573836Vilma Torres GarcíaAún no hay calificaciones
- Aparato digestivo: desarrollo y formación de órganosDocumento6 páginasAparato digestivo: desarrollo y formación de órganosLeodan Antonio Macas GonzalezAún no hay calificaciones
- CUESTIONARIO Derivados EndodermalesDocumento5 páginasCUESTIONARIO Derivados Endodermalesapi-3722643100% (1)
- Desarrollo Del Intestino AnteriorDocumento4 páginasDesarrollo Del Intestino AnteriorErikaAún no hay calificaciones
- Desarrollo EmbrionarioDocumento6 páginasDesarrollo EmbrionariomelisaAún no hay calificaciones
- Sistema DigestivoDocumento2 páginasSistema DigestivoJoseph FaríasAún no hay calificaciones
- Embrio Sistema Digestivo (Resumen)Documento8 páginasEmbrio Sistema Digestivo (Resumen)1er año MED.Aún no hay calificaciones
- Práctica 6 Aparato Digestivo y Glándulas AnexasDocumento8 páginasPráctica 6 Aparato Digestivo y Glándulas AnexasShelsie Suarez GarciaAún no hay calificaciones
- Cuestionario Sobre Los Aparatos Respiratorio y Digestivo EmbriologiaDocumento4 páginasCuestionario Sobre Los Aparatos Respiratorio y Digestivo EmbriologiaDavid FloresAún no hay calificaciones
- Embriologia Del Sistema DigestivoDocumento18 páginasEmbriologia Del Sistema DigestivoDomingo Sanoja GimonAún no hay calificaciones
- 15 DigestivoDocumento18 páginas15 DigestivoSebas YMAún no hay calificaciones
- Sistema Digestivo Embriologia 233676 Downloable 940677Documento10 páginasSistema Digestivo Embriologia 233676 Downloable 940677Kenat Madai L�pez Ju�rezAún no hay calificaciones
- Desarrollo Embrionario Del Aparato DigestivoDocumento38 páginasDesarrollo Embrionario Del Aparato DigestivoFiorella LrAún no hay calificaciones
- Capítulo 13. Desarrollo Del Sistema DigestivoDocumento10 páginasCapítulo 13. Desarrollo Del Sistema DigestivoIgnacio PerezAún no hay calificaciones
- Clase 3 y 4 Embriologia Iii Ciclo 2021 - IiDocumento43 páginasClase 3 y 4 Embriologia Iii Ciclo 2021 - IiTania CAún no hay calificaciones
- Sistema Digestivo - Documentos de GoogleDocumento9 páginasSistema Digestivo - Documentos de GoogleKristel Urbina CascoAún no hay calificaciones
- GlosariosDocumento29 páginasGlosariosMayrisAún no hay calificaciones
- Aparato Digestivo HistoDocumento7 páginasAparato Digestivo HistoPlay StoryAún no hay calificaciones
- Malformaciones digestivas: diagnóstico y tratamientoDocumento65 páginasMalformaciones digestivas: diagnóstico y tratamientoIsabella CastilloAún no hay calificaciones
- 3.7-3.9 EmbriologiaDocumento10 páginas3.7-3.9 EmbriologiaDiego GervacioAún no hay calificaciones
- ESTOMAGODocumento6 páginasESTOMAGOHugo MiguelAún no hay calificaciones
- Desarrollo de Tubo DigestivoDocumento6 páginasDesarrollo de Tubo Digestivoyviggu100% (1)
- Desarrollo Del Aparato DigestivoDocumento17 páginasDesarrollo Del Aparato DigestivoAna Julia Domingues CrivelliAún no hay calificaciones
- Aparato Digestivo (TP 8)Documento5 páginasAparato Digestivo (TP 8)Araceli RetamozoAún no hay calificaciones
- Revisià N Embriologà A Abdominal Juan Pablo Borrayo EscobarDocumento11 páginasRevisià N Embriologà A Abdominal Juan Pablo Borrayo EscobarrcastacAún no hay calificaciones
- Capitulo 15 Sistema DigestivoDocumento6 páginasCapitulo 15 Sistema DigestivoRachell CaceresAún no hay calificaciones
- Presentación1 SISTEMA DIGESTIVODocumento60 páginasPresentación1 SISTEMA DIGESTIVODennis VasquezAún no hay calificaciones
- (Spanish (Auto-Generated) ) EMBRIOLOGÍA Del APARATO DIGESTIVO ?? (DownSub - Com)Documento8 páginas(Spanish (Auto-Generated) ) EMBRIOLOGÍA Del APARATO DIGESTIVO ?? (DownSub - Com)Catalina boscanAún no hay calificaciones
- Sem 10 Práctica Aparato Digestivo I.dr GonzalesDocumento23 páginasSem 10 Práctica Aparato Digestivo I.dr GonzalesNicolle Aquino ThorneAún no hay calificaciones
- Sistema DigestivoDocumento42 páginasSistema DigestivoJUAN ENRIQUE CORTEZ QUIROGAAún no hay calificaciones
- Desarrollo del intestino primitivo y órganos anexosDocumento11 páginasDesarrollo del intestino primitivo y órganos anexosGj3len :3Aún no hay calificaciones
- Embriología del sistema digestivo enDocumento20 páginasEmbriología del sistema digestivo enCristianAún no hay calificaciones
- Embarazo, parto y lactancia del bebéDe EverandEmbarazo, parto y lactancia del bebéAún no hay calificaciones
- Efectos de La TecnologiaDocumento7 páginasEfectos de La TecnologiaMallma ProyectosAún no hay calificaciones
- Efectos de La TecnologiaDocumento7 páginasEfectos de La TecnologiaMallma ProyectosAún no hay calificaciones
- Queso FrescoDocumento11 páginasQueso FrescoMallma Proyectos100% (1)
- Efectos de La TecnologiaDocumento7 páginasEfectos de La TecnologiaMallma ProyectosAún no hay calificaciones
- Presentacion LuuDocumento14 páginasPresentacion LuuMallma ProyectosAún no hay calificaciones
- Modelo de Acto Constitutivo de Sociedad Comercial de Responsabilidad Limitada 1Documento3 páginasModelo de Acto Constitutivo de Sociedad Comercial de Responsabilidad Limitada 1Mallma ProyectosAún no hay calificaciones
- Infome Pozo A TierraDocumento10 páginasInfome Pozo A TierraMallma ProyectosAún no hay calificaciones
- Breve Historia de EspaniaDocumento7 páginasBreve Historia de EspanianahoriAún no hay calificaciones
- Etica 2012 Clase 1 DefDocumento38 páginasEtica 2012 Clase 1 DefMallma ProyectosAún no hay calificaciones
- Tesis EjemploDocumento75 páginasTesis Ejemplojuan jose soto cahuaAún no hay calificaciones
- Enfermedades Transmitidas Por PrionesDocumento6 páginasEnfermedades Transmitidas Por PrionesMallma ProyectosAún no hay calificaciones
- Zoonosis Emergentes y Reemergentes-3Documento7 páginasZoonosis Emergentes y Reemergentes-3Jimmy GAAún no hay calificaciones
- 01 Cursoderedes 02Documento72 páginas01 Cursoderedes 02Franklin Quispe Arpi100% (1)
- Etica 2012 Clase 1 DefDocumento38 páginasEtica 2012 Clase 1 DefMallma ProyectosAún no hay calificaciones
- Como Escribir Una Tesis - Umberto EcoDocumento51 páginasComo Escribir Una Tesis - Umberto EcoCECADI29100% (1)
- Tutorial Escaner OBD2 PDFDocumento182 páginasTutorial Escaner OBD2 PDFHomer Sadych Henao MarinAún no hay calificaciones
- Neuma TicaDocumento10 páginasNeuma TicaMallma ProyectosAún no hay calificaciones
- Obd IiDocumento32 páginasObd Iicamwsa100% (2)
- RVM N 064 2018 Minedu BasesDocumento2 páginasRVM N 064 2018 Minedu BasesMallma ProyectosAún no hay calificaciones
- 1-Manual Diesel Pesados Mercedes Benz PLDDocumento22 páginas1-Manual Diesel Pesados Mercedes Benz PLDinyecciolanave94% (210)
- Manual de NeumaticaDocumento281 páginasManual de Neumaticacordobaweb3688100% (4)
- PlanDocumento29 páginasPlanMallma ProyectosAún no hay calificaciones
- MUSCULOS Torácicos AnterioresDocumento8 páginasMUSCULOS Torácicos AnterioresCarlos CamargoAún no hay calificaciones
- Examen Control AnatomíaDocumento8 páginasExamen Control AnatomíaIsaíasGermánRomeroQuicioAún no hay calificaciones
- Laringe Intro VozDocumento5 páginasLaringe Intro VozMili SkafAún no hay calificaciones
- Volcanes de GuatemalaDocumento7 páginasVolcanes de Guatemalabrenda cac xolAún no hay calificaciones
- MICCIÒNDocumento4 páginasMICCIÒNCalep OrdoñezAún no hay calificaciones
- Epistaxis en LeishmaniaDocumento4 páginasEpistaxis en Leishmaniasoledad100% (1)
- Microbiota emocionesDocumento4 páginasMicrobiota emocionesLudita Vento100% (1)
- TIPOS DE CONTRACCIONES MuscularesDocumento13 páginasTIPOS DE CONTRACCIONES MuscularesAntoine VI/ROAún no hay calificaciones
- Anatomia de Pelvis OseaDocumento16 páginasAnatomia de Pelvis OseaMariana Ramos GarciaAún no hay calificaciones
- Citología ExfoliativaDocumento1 páginaCitología ExfoliativaRegina AlpizarAún no hay calificaciones
- Neurotransmisores y ClasificacionDocumento59 páginasNeurotransmisores y ClasificacionCeci Flores100% (1)
- Caso de Estudio 1 PatologiaDocumento4 páginasCaso de Estudio 1 PatologiaAndrea VargasAún no hay calificaciones
- Modelo 6: Visceras abdominales optimizadas paraDocumento17 páginasModelo 6: Visceras abdominales optimizadas paraSamuel AjpacajaAún no hay calificaciones
- GUIA DE ANATOMÍA SESIÓN 13 Aparato Urinario 2Documento9 páginasGUIA DE ANATOMÍA SESIÓN 13 Aparato Urinario 2JULISSA MERCEDES CORTEZ COKCHIAún no hay calificaciones
- EL SISTEMA NERVIOSO 2 GradoDocumento6 páginasEL SISTEMA NERVIOSO 2 Gradomartha suarezAún no hay calificaciones
- Notas Pato 2Documento32 páginasNotas Pato 2Leilani HerreraAún no hay calificaciones
- Sistema Nervioso AutónomoDocumento10 páginasSistema Nervioso AutónomoLilymar MorenoAún no hay calificaciones
- Procedimientos Respiratorios PDFDocumento335 páginasProcedimientos Respiratorios PDFAbel NetoAún no hay calificaciones
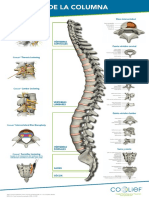
- Anatomia de La Columna VertebralDocumento1 páginaAnatomia de La Columna VertebralAngel Fernandez100% (1)
- Sentidos CaballosDocumento10 páginasSentidos CaballosrosersalouAún no hay calificaciones
- Examen Final de Gineco ObstetriciaDocumento5 páginasExamen Final de Gineco ObstetriciaLennin Alcántara ValleAún no hay calificaciones
- Guia Digital Masajes Terapeuticos PracticoDocumento42 páginasGuia Digital Masajes Terapeuticos PracticoMarlyn AafAún no hay calificaciones
- TRAQUEADocumento2 páginasTRAQUEArdi09Aún no hay calificaciones
- Tejido OseoDocumento2 páginasTejido OseoroseAún no hay calificaciones
- Proyecciones Básicas y Específicas para CráneoDocumento8 páginasProyecciones Básicas y Específicas para Cráneosuperloki25Aún no hay calificaciones
- Perfil TiroideoDocumento16 páginasPerfil TiroideofernandaAún no hay calificaciones
- Fisiopatología de La Tuberculosis: Wilmer Guzmán Ventura Internista - NefrólogoDocumento22 páginasFisiopatología de La Tuberculosis: Wilmer Guzmán Ventura Internista - NefrólogoMax100% (1)
- CerdosDocumento8 páginasCerdosCristian Exequiel CarbajalAún no hay calificaciones
- RETINADocumento25 páginasRETINAGuillermo RomeroAún no hay calificaciones
- Anatomia AviarDocumento2 páginasAnatomia Aviarnorma paulina carcausto lipaAún no hay calificaciones