También podría gustarte
- Tema 11Documento4 páginasTema 11Miguel Navarro ViñualesAún no hay calificaciones
- Animales InvertebradosDocumento7 páginasAnimales InvertebradosDiana Parrado CAún no hay calificaciones
- Fisiologia Reproductiva en La Hembra de AnfibiosDocumento8 páginasFisiologia Reproductiva en La Hembra de AnfibiosANAMARIAAún no hay calificaciones
- Reproduccion en MoluscosDocumento15 páginasReproduccion en MoluscosfiorelaAún no hay calificaciones
- EquinodermosDocumento9 páginasEquinodermosAndrea MedeloAún no hay calificaciones
- Morfología de GónadasDocumento4 páginasMorfología de Gónadasanon_864211228Aún no hay calificaciones
- BG03BAZÁN-Reproducción Animal INVERTEDocumento2 páginasBG03BAZÁN-Reproducción Animal INVERTEAnonymous 7za15o0Aún no hay calificaciones
- Reproduccion AnimalDocumento6 páginasReproduccion AnimalTulibeth Maestre Gonzalez100% (1)
- Desarrollo Temprano de Los CaracolesDocumento6 páginasDesarrollo Temprano de Los CaracolesMario Lopez JumboAún no hay calificaciones
- SM Función de Reproducción en AnimalesDocumento37 páginasSM Función de Reproducción en AnimalesAlexander Medina VolkovAún no hay calificaciones
- La Embriología de Los Animales AcuáticosDocumento6 páginasLa Embriología de Los Animales AcuáticosErick Rodrigo Valdiviezo PantaAún no hay calificaciones
- Unidad 7. La Reproducciã - N en Los AnimalesDocumento19 páginasUnidad 7. La Reproducciã - N en Los Animalesimane erritaliAún no hay calificaciones
- Ciclo Biológico de InsectosDocumento30 páginasCiclo Biológico de InsectosClaudia Vera QuilcaAún no hay calificaciones
- Desarrollo EmbrionarioDocumento20 páginasDesarrollo EmbrionarioAMILCYRAún no hay calificaciones
- ANFIOXUSDocumento7 páginasANFIOXUSTJJeffersonAún no hay calificaciones
- Diferentes Tipos de ReproduccionDocumento27 páginasDiferentes Tipos de ReproduccionAlejandro MuñozAún no hay calificaciones
- Taller #2 ZoologiaDocumento22 páginasTaller #2 ZoologiaAndres Camilo Velez HerreraAún no hay calificaciones
- Bloque Temático 2 - T5Documento10 páginasBloque Temático 2 - T5Álvarito DelgadoAún no hay calificaciones
- DESARROLLO EMBRIONARIO EN ERIZO DE MAR Tetrapygus Niger - AsdDocumento8 páginasDESARROLLO EMBRIONARIO EN ERIZO DE MAR Tetrapygus Niger - AsdYanina MadeleiiAún no hay calificaciones
- Apunte de Sistema Reproductor en Vertebrados e InvertebradosDocumento44 páginasApunte de Sistema Reproductor en Vertebrados e Invertebradoslu45zxcv50% (2)
- Clase 02 CaracterizacionDocumento36 páginasClase 02 CaracterizacionRocio Elizabeth ChaptuAún no hay calificaciones
- Embriogenesis de Los AnfibiosDocumento4 páginasEmbriogenesis de Los AnfibiosFabiAún no hay calificaciones
- OSBGT45Documento13 páginasOSBGT45alexantonjazzAún no hay calificaciones
- Diversidad Animal 2Documento14 páginasDiversidad Animal 2Diana Judith TorresAún no hay calificaciones
- Desarrollo EmbrionarioDocumento13 páginasDesarrollo EmbrionarioMaria ChavielAún no hay calificaciones
- Animales Vertebrados e InvertebradosDocumento13 páginasAnimales Vertebrados e InvertebradosVictor Daniel Flores CoapazaAún no hay calificaciones
- Tipos y Características de Huevos Desde Peces Hasta MamíferosDocumento8 páginasTipos y Características de Huevos Desde Peces Hasta MamíferosJazmin Maritza Benito MamaniAún no hay calificaciones
- Guia Reproducción en AnimalesDocumento3 páginasGuia Reproducción en AnimalesLeider Mauricio Valencia RubianoAún no hay calificaciones
- Desarrollo Embrionario de Los AnfibiosDocumento4 páginasDesarrollo Embrionario de Los AnfibiosJavier LugoAún no hay calificaciones
- Cuestionario Desarrollo de PecesDocumento2 páginasCuestionario Desarrollo de PecesEvelynAún no hay calificaciones
- Reproduccion de ReptilesDocumento21 páginasReproduccion de ReptilesPliner Morales RAún no hay calificaciones
- Sistema Reproductor de Reptiles y PecesDocumento4 páginasSistema Reproductor de Reptiles y PecesAldo FernandezAún no hay calificaciones
- Informepeces y AnfibiosDocumento16 páginasInformepeces y AnfibiosLORENS NIKOLL SANCHEZ MARTINEZAún no hay calificaciones
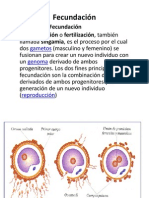
- FecundaciónDocumento3 páginasFecundaciónJuli lopezAún no hay calificaciones
- Histología de Gónadas y Caracterización de Gametos en Peces y AnfibiosDocumento4 páginasHistología de Gónadas y Caracterización de Gametos en Peces y AnfibiosChristian Moises CasasAún no hay calificaciones
- ALGASDocumento5 páginasALGAS.:("*"BLacK BuLLeT"*"):.0% (1)
- RecurrenDocumento3 páginasRecurrenMiguel JimenezAún no hay calificaciones
- Tipos de HuevosDocumento23 páginasTipos de Huevoskarin1883% (6)
- GasteropodosDocumento6 páginasGasteropodosJorgue Leonardo RodriguezAún no hay calificaciones
- 5Documento12 páginas5Carlos AlcaAún no hay calificaciones
- Presentacion FecundacionDocumento63 páginasPresentacion FecundacionMelissa CórdobaAún no hay calificaciones
- Sem 7 Desarrollo Embrionario 01jul2019Documento43 páginasSem 7 Desarrollo Embrionario 01jul2019Jose Luis Urbina ChorresAún no hay calificaciones
- Los Opalinidos Son Protozoarios Multinucleados o BinucleadosDocumento2 páginasLos Opalinidos Son Protozoarios Multinucleados o BinucleadosDanne Mendoza100% (1)
- ZARIGUELLASDocumento14 páginasZARIGUELLASLina Julieth Mendez CardenasAún no hay calificaciones
- BIOLOGÍA Y GEOLOGÍA Tema 16. Reproducción en AnimalesDocumento27 páginasBIOLOGÍA Y GEOLOGÍA Tema 16. Reproducción en AnimalesMaría José Morales100% (1)
- Animales Sexualesn y AsexualesDocumento2 páginasAnimales Sexualesn y AsexualesLiliana Patricia Olivares OlivaresAún no hay calificaciones
- Informe Final 1 BDDocumento12 páginasInforme Final 1 BDLOPEZ ACOSTA INGRITH NATALIAAún no hay calificaciones
- 1.8c Reproducción ArtrópodosDocumento7 páginas1.8c Reproducción ArtrópodosYahir Emanuel Reyes BalamAún no hay calificaciones
- Reproduccion en AnimalesDocumento44 páginasReproduccion en AnimalesMariana León100% (1)
- 2-Reproducc Ion Sexual-1Documento9 páginas2-Reproducc Ion Sexual-1Luna CruzAún no hay calificaciones
- ReproduccionanimalDocumento24 páginasReproduccionanimalLukas LazarenoAún no hay calificaciones
- Desove y Fecundacion de CrustaceosDocumento10 páginasDesove y Fecundacion de Crustaceosmilton clavel magiano shavitAún no hay calificaciones
- Tipos de FecundaciónDocumento5 páginasTipos de Fecundaciónamandanet5Aún no hay calificaciones
- Reproduccion y Defensa en Invertebrados - FinalDocumento44 páginasReproduccion y Defensa en Invertebrados - Finalana jenci restrepoAún no hay calificaciones
- Resumen de El Proceso de Hominización: RESÚMENES UNIVERSITARIOSDe EverandResumen de El Proceso de Hominización: RESÚMENES UNIVERSITARIOSAún no hay calificaciones
- Anatomía artística 9: Mamíferos: morfología comparadaDe EverandAnatomía artística 9: Mamíferos: morfología comparadaAún no hay calificaciones
- Plantilla Hoja de PresentacionDocumento1 páginaPlantilla Hoja de PresentacionMarie AnAún no hay calificaciones
- Crinoideos o Lirios de MarDocumento4 páginasCrinoideos o Lirios de MarMarie AnAún no hay calificaciones
- Reproduccion y Desarrollo (De La Web)Documento9 páginasReproduccion y Desarrollo (De La Web)Marie AnAún no hay calificaciones
- DensidadDocumento2 páginasDensidadEtni manuela Rivera MataAún no hay calificaciones
- Ejercicios DinámicaDocumento4 páginasEjercicios DinámicaMarie AnAún no hay calificaciones
- Ejercicios DinámicaDocumento4 páginasEjercicios DinámicaMarie AnAún no hay calificaciones
- Práctica - Energía. FísicaDocumento2 páginasPráctica - Energía. FísicaMarie AnAún no hay calificaciones
- Números ComplejosDocumento4 páginasNúmeros ComplejosMarie AnAún no hay calificaciones
- FORMA POLAR DE NÚMEROS COMPLEJOS PagDocumento4 páginasFORMA POLAR DE NÚMEROS COMPLEJOS PagMarie AnAún no hay calificaciones
- TGS - PC2 - Sanchez - Contreras - Edwart PatherssonDocumento8 páginasTGS - PC2 - Sanchez - Contreras - Edwart PatherssonEdwart SanchezAún no hay calificaciones
- Tema 1 Gral Ant Sistemica y TPGFC IDocumento19 páginasTema 1 Gral Ant Sistemica y TPGFC IKeydellAún no hay calificaciones
- Glosario Etimológico de MariposasDocumento62 páginasGlosario Etimológico de MariposasHANDREY100% (1)
- JirafaDocumento4 páginasJirafaDariela Ureña diazAún no hay calificaciones
- Etologia C.socialDocumento4 páginasEtologia C.socialConcepcion Sullon ChanduviAún no hay calificaciones
- 14 La BiorregeneracionDocumento70 páginas14 La BiorregeneracionSebastián Reina CanoAún no hay calificaciones
- Potencial GeneradorDocumento1 páginaPotencial GeneradorPablo Vega MAún no hay calificaciones
- Dieta Currucuti Edellin PDFDocumento4 páginasDieta Currucuti Edellin PDFLeslie CadavidAún no hay calificaciones
- Ab Iyo Ed 09 218 00Documento50 páginasAb Iyo Ed 09 218 00Walter EnriqueAún no hay calificaciones
- 01 Embriologia y Anatomia OcularDocumento50 páginas01 Embriologia y Anatomia OcularluisAún no hay calificaciones
- Caja ToracicaDocumento15 páginasCaja ToracicaPaulina Almeida RiccettoAún no hay calificaciones
- Biol emDocumento99 páginasBiol emJoel VegaAún no hay calificaciones
- Ensayo Sobre ImportanciaDocumento2 páginasEnsayo Sobre ImportanciadesireAún no hay calificaciones
- La " Hora de Juego Diagnóstica", Es Entendida Por Alicia Fernández, Como: "Este Espacio de Encuentro Entre El Profesional y El Niño, Supera LaDocumento15 páginasLa " Hora de Juego Diagnóstica", Es Entendida Por Alicia Fernández, Como: "Este Espacio de Encuentro Entre El Profesional y El Niño, Supera LaRocio CarbajalAún no hay calificaciones
- NATURALES Los Hongos Del 7 Al 25 de SeptDocumento4 páginasNATURALES Los Hongos Del 7 Al 25 de SeptMariselly BustosAún no hay calificaciones
- El Sustantivo Individual y ColectivoDocumento6 páginasEl Sustantivo Individual y ColectivoErika Natali Tantalean MesonesAún no hay calificaciones
- Examen Resuelto MicrobiologiaDocumento2 páginasExamen Resuelto MicrobiologiaDarwin VargasAún no hay calificaciones
- 5.3.7 RevegetaciónDocumento42 páginas5.3.7 RevegetaciónCESARAún no hay calificaciones
- WildDocumento208 páginasWildSelena GuerraAún no hay calificaciones
- Guía N°1 El Almohadon de PlumasDocumento5 páginasGuía N°1 El Almohadon de PlumasInambu Lp100% (1)
- Clase 1 Genetica 148Documento48 páginasClase 1 Genetica 148scottAún no hay calificaciones
- Cuaderno de Griego IDocumento114 páginasCuaderno de Griego IVioletta67Aún no hay calificaciones
- Glosario BiologiaDocumento2 páginasGlosario BiologiaJimena GuerreroAún no hay calificaciones
- PUD 7mo.Documento5 páginasPUD 7mo.Florencio AbadAún no hay calificaciones
- Sistema Nervioso U2-1 - SN Estìmulos-Respuestas - Clasificaciòn - Estructura y Función - 20222Documento3 páginasSistema Nervioso U2-1 - SN Estìmulos-Respuestas - Clasificaciòn - Estructura y Función - 20222nayleth zamarik camacho carrilloAún no hay calificaciones
- Cromosomas GigantesDocumento3 páginasCromosomas GigantesRicardo TorresAún no hay calificaciones
- Ej 1 Ser VivoDocumento3 páginasEj 1 Ser VivoRenato CoroAún no hay calificaciones
- 1.2. Unidad 1 OKDocumento39 páginas1.2. Unidad 1 OKCiclay Cagua100% (1)
- T1 BiologiaDocumento12 páginasT1 BiologiaJose Angel Dimas SalinasAún no hay calificaciones
- Biología CitologiaDocumento5 páginasBiología CitologiaLlanos Durán Jafet EsmeraldaAún no hay calificaciones