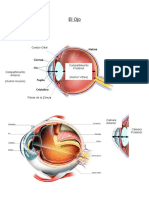
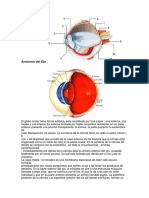
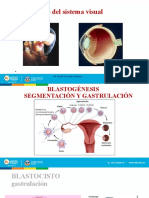
También podría gustarte
- Beneficios del Cannabis Medicinal en el GlaucomaDe EverandBeneficios del Cannabis Medicinal en el GlaucomaAún no hay calificaciones
- Embriologia OcularDocumento20 páginasEmbriologia OcularEstephany SantosAún no hay calificaciones
- Embriologia Del Globo OcularDocumento11 páginasEmbriologia Del Globo OcularKaroline Santana100% (2)
- Informe Ojo y Oido HistologíaDocumento38 páginasInforme Ojo y Oido HistologíaJosé LuisAún no hay calificaciones
- Curso de Iluminación Indalux CompletoDocumento264 páginasCurso de Iluminación Indalux Completoalucard 019Aún no hay calificaciones
- La Ciencia de La Longevidad PDFDocumento99 páginasLa Ciencia de La Longevidad PDFJose Ramon100% (3)
- Desarrollo Embrionario Del OjoDocumento2 páginasDesarrollo Embrionario Del OjoCarlosSonoAún no hay calificaciones
- Embriología OcularDocumento49 páginasEmbriología OcularLigia Rios100% (1)
- Desarrollo Del OjoDocumento23 páginasDesarrollo Del OjoStiven GarcíaAún no hay calificaciones
- Embriología Del Ojo - VALERIANODocumento6 páginasEmbriología Del Ojo - VALERIANOJosue ValerianoAún no hay calificaciones
- Embriogenesis Del Ojo 2014Documento52 páginasEmbriogenesis Del Ojo 2014WalterQunoqYanceOw100% (1)
- Retina - Corregida - PPTX Filename - UTF-8''Retina CorregidaDocumento97 páginasRetina - Corregida - PPTX Filename - UTF-8''Retina CorregidaLMN214Aún no hay calificaciones
- Embriología Del Globo OcularDocumento8 páginasEmbriología Del Globo OcularYorgenis Nuñez100% (1)
- Embriologia Del OjoDocumento37 páginasEmbriologia Del OjoMilena VeraAún no hay calificaciones
- Anatomía Del Globo Ocular Prac de Oftalmologia AnalDocumento18 páginasAnatomía Del Globo Ocular Prac de Oftalmologia AnalWelinson Quezada100% (1)
- Embriologia OcularDocumento49 páginasEmbriologia OcularEsther AlonsoAún no hay calificaciones
- Embriología Del OjoDocumento8 páginasEmbriología Del OjoPatrick NovoaAún no hay calificaciones
- Embriología Del OjoDocumento9 páginasEmbriología Del OjoBruno FernandoAún no hay calificaciones
- Embriologia RetinaDocumento6 páginasEmbriologia RetinaedenmarrugoAún no hay calificaciones
- Ojo y OidoDocumento5 páginasOjo y OidoYeltsin DaynorAún no hay calificaciones
- Tema XXI Desarrollo de Ojos y OidosDocumento9 páginasTema XXI Desarrollo de Ojos y OidosKairich V SojoAún no hay calificaciones
- Compendio de Embriología OcularDocumento34 páginasCompendio de Embriología OcularFelipe NaranjoAún no hay calificaciones
- Reseña Embriológica y de La Estructura Del Aparato Visual, Sus Principales Malformaciones CongénitasDocumento23 páginasReseña Embriológica y de La Estructura Del Aparato Visual, Sus Principales Malformaciones CongénitasJosé Gabriel Quiroga Villagra100% (1)
- Fisiologia de La Visión PDFDocumento30 páginasFisiologia de La Visión PDFMédecin Adrian TGAún no hay calificaciones
- Embriología Del Globo Ocular.Documento13 páginasEmbriología Del Globo Ocular.Eduardo EstrellaAún no hay calificaciones
- Organo de La VisionDocumento33 páginasOrgano de La Visionjunior pinedaAún no hay calificaciones
- Cuestionario Ojo OidoDocumento10 páginasCuestionario Ojo OidoPaula EloinaAún no hay calificaciones
- Informe de Ojo y OidoDocumento15 páginasInforme de Ojo y OidoKarenVivianaAún no hay calificaciones
- Órgano de Los SentidosDocumento55 páginasÓrgano de Los SentidosUSMP FN ARCHIVOSAún no hay calificaciones
- Embriología Del Globo OcularDocumento7 páginasEmbriología Del Globo OcularKarla LeónAún no hay calificaciones
- 15 Embriologia Ojo y Oido.Documento9 páginas15 Embriologia Ojo y Oido.Yonger Leandro Guilarte DuranAún no hay calificaciones
- INTRODUCCIÓNDocumento20 páginasINTRODUCCIÓNLorenzo QuintanaAún no hay calificaciones
- Ojo y OídoDocumento11 páginasOjo y OídoSamantha PonceAún no hay calificaciones
- Resumen OjoDocumento11 páginasResumen OjomaríaAún no hay calificaciones
- Anatomía Del OjoDocumento22 páginasAnatomía Del Ojoyargmilin mendozaAún no hay calificaciones
- Diapos Organos de Los SentidosDocumento11 páginasDiapos Organos de Los SentidosAna Valeria Casiano CelisAún no hay calificaciones
- Seminario Desarrollo Embrionario Del Ojo 1Documento21 páginasSeminario Desarrollo Embrionario Del Ojo 1Hermes Carruyo MoranAún no hay calificaciones
- Abp de Vide para MorfologiaDocumento10 páginasAbp de Vide para Morfologiadanna belloAún no hay calificaciones
- Desarrollo de Los Órganos de Los SentidosDocumento6 páginasDesarrollo de Los Órganos de Los SentidosRosmary HornaAún no hay calificaciones
- Anatomía y Fisiología Del OjoDocumento26 páginasAnatomía y Fisiología Del Ojowilybarrera100% (1)
- Desarrollo Del OidoDocumento5 páginasDesarrollo Del Oidolarapaloma698Aún no hay calificaciones
- Embriologia Del OjoDocumento1 páginaEmbriologia Del OjoJenny SarzosaAún no hay calificaciones
- Desarrollo Ojo y OidoDocumento9 páginasDesarrollo Ojo y OidoSofia PujalteAún no hay calificaciones
- Presentacion EmbriogenesisDocumento31 páginasPresentacion EmbriogenesisJey P. AchaiahAún no hay calificaciones
- Informe Embrio-Microb Seccion U 4TA PromoDocumento9 páginasInforme Embrio-Microb Seccion U 4TA PromoJose Turcios EucedaAún no hay calificaciones
- Embriologia Tercer Parcial OjoDocumento36 páginasEmbriologia Tercer Parcial OjoRay FunesAún no hay calificaciones
- Informe de EmbriologiaDocumento17 páginasInforme de EmbriologiajoseAún no hay calificaciones
- Anatomía Del Globo Ocular Prac de Oftalmologia Analia Rosa 1-18-0556...Documento18 páginasAnatomía Del Globo Ocular Prac de Oftalmologia Analia Rosa 1-18-0556...Welinson QuezadaAún no hay calificaciones
- Resumen de Ojo EmbriologíaDocumento3 páginasResumen de Ojo EmbriologíaJuan CAún no hay calificaciones
- Clase 1 Retina ADocumento6 páginasClase 1 Retina AEma Antonia AlmarzaAún no hay calificaciones
- Embriologia Del Ojo Final.Documento19 páginasEmbriologia Del Ojo Final.Camilo Andres Moreno ChicaAún no hay calificaciones
- Ojo Seminario de EmbriologiaDocumento6 páginasOjo Seminario de EmbriologiaCinthia SamaméAún no hay calificaciones
- 2015 Estesiologia (Ojo y Oido)Documento19 páginas2015 Estesiologia (Ojo y Oido)Franco MatusAún no hay calificaciones
- Embriología OcularDocumento6 páginasEmbriología OcularMaría Ángeles NeiraAún no hay calificaciones
- Neuroanatomía Embriología Del Ojo DiapositivasDocumento25 páginasNeuroanatomía Embriología Del Ojo DiapositivasElizabeth Saenz PazmiñoAún no hay calificaciones
- Teoria de OftamologiaDocumento8 páginasTeoria de OftamologiaCapitan BarbosaAún no hay calificaciones
- Ojo y Oído-UnprgDocumento56 páginasOjo y Oído-UnprgS Daphne BautistaAún no hay calificaciones
- Embriología y Anatomía Del OjoDocumento17 páginasEmbriología y Anatomía Del OjoCarmen Sofia Mejia PayvaAún no hay calificaciones
- Embriología OcularDocumento15 páginasEmbriología OcularFrancisco Palma CarvajalAún no hay calificaciones
- Embriologia Retina, No, UveaDocumento3 páginasEmbriologia Retina, No, UveaMARIANAún no hay calificaciones
- EMBRIOLOGIA OCULAR, Tercer Semestre-1-48Documento48 páginasEMBRIOLOGIA OCULAR, Tercer Semestre-1-48MAGNOLIA ARANGOAún no hay calificaciones
- Embriologia 2022 PDFDocumento17 páginasEmbriologia 2022 PDFWillians Cabrera FleitasAún no hay calificaciones
- Estructura Del OjoDocumento3 páginasEstructura Del Ojomaricute851Aún no hay calificaciones
- Tema 21. Oftalmología PediatricaDocumento83 páginasTema 21. Oftalmología PediatricapaulfjsdAún no hay calificaciones
- Tecnicas de IluminacionDocumento2 páginasTecnicas de Iluminacionandrea saavedraAún no hay calificaciones
- Guía Docente de La Asignatura Principios de OptometríaDocumento12 páginasGuía Docente de La Asignatura Principios de OptometríaDorian BallasAún no hay calificaciones
- 1.2 Confort VisualDocumento12 páginas1.2 Confort Visuallucia veraAún no hay calificaciones
- Cabeza Y Cuello: Aporte: Mg. Hugo Cerdán CuevaDocumento113 páginasCabeza Y Cuello: Aporte: Mg. Hugo Cerdán CuevaJohnattan Javier Espinoza CastañedaAún no hay calificaciones
- MOLADA - Tratamiento de Datos Colorimétricos y Espectrales en Documentación de Arte Rupestre. El ...Documento161 páginasMOLADA - Tratamiento de Datos Colorimétricos y Espectrales en Documentación de Arte Rupestre. El ...JUAN CAMILO GOMEZ QUINTEROAún no hay calificaciones
- Oftalmologia Resumen 2004-2005Documento6 páginasOftalmologia Resumen 2004-2005KATTY2125Aún no hay calificaciones
- Trabajo OftalmologiaDocumento26 páginasTrabajo Oftalmologialuisgregori acuñaAún no hay calificaciones
- Lineas de InvestigacionDocumento4 páginasLineas de InvestigacionfeAún no hay calificaciones
- Traumatismo OcularDocumento38 páginasTraumatismo OcularBryan MontoyaAún no hay calificaciones
- Existen Dos Tipos de Lentes Progresivas: Convencionales y DigitalesDocumento7 páginasExisten Dos Tipos de Lentes Progresivas: Convencionales y DigitalesAlejo CedeñoAún no hay calificaciones
- Cómo Ven Los AnimalesDocumento3 páginasCómo Ven Los AnimalesEli KimAún no hay calificaciones
- Piel y Organos de Los SentidosDocumento66 páginasPiel y Organos de Los SentidosLaura TorresAún no hay calificaciones
- Carta de Snellen para Determinacion de Agudeza VisualDocumento4 páginasCarta de Snellen para Determinacion de Agudeza VisualArlette Herrera0% (1)
- TESINA - Desarrollo de Un Helado Reducido en Lactosa - MEDINA - INESDocumento68 páginasTESINA - Desarrollo de Un Helado Reducido en Lactosa - MEDINA - INESMark AlexanderAún no hay calificaciones
- Oftarmologia Doctor Cesar BeleDocumento43 páginasOftarmologia Doctor Cesar Bele100470747nasmAún no hay calificaciones
- Mapas MentalesDocumento17 páginasMapas Mentalesgloria cardenasAún no hay calificaciones
- Semiologia Neurologica 01Documento84 páginasSemiologia Neurologica 01Anderson fox liveAún no hay calificaciones
- Introducción A La Anatomía y Fisiología OcularDocumento13 páginasIntroducción A La Anatomía y Fisiología OcularYuli Orti´zAún no hay calificaciones
- Of 16 2 002Documento22 páginasOf 16 2 002David MartínezAún no hay calificaciones
- Prueba Cnat. Los Sentidos 3º PrimariaDocumento3 páginasPrueba Cnat. Los Sentidos 3º PrimariaRosa Esther Mora Espino100% (1)
- Estudio Arancelario - ComercioDocumento154 páginasEstudio Arancelario - ComercioEvelin Yulieth Castro GraciaAún no hay calificaciones
- Aberración CromáticaDocumento3 páginasAberración CromáticaVilcazan Huayta JuanAún no hay calificaciones
- Tema24 Resumen Desprendimiento de RetinaDocumento3 páginasTema24 Resumen Desprendimiento de Retinamayela de la rosaAún no hay calificaciones
- Guía Clínica de GlaucomaDocumento5 páginasGuía Clínica de GlaucomaMaritza VelardeAún no hay calificaciones
- 1 Fisica RefraccionDocumento8 páginas1 Fisica RefraccionPatricio Eugenio Hinrichsen LippiAún no hay calificaciones