También podría gustarte
- Anatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2De EverandAnatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2Aún no hay calificaciones
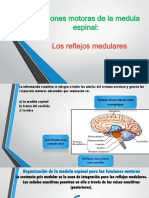
- Funciones Motoras de La Medula Espinal Reflejos Medulares DR MerelloDocumento83 páginasFunciones Motoras de La Medula Espinal Reflejos Medulares DR Merellojosemerello100% (1)
- Anatomía y Fisiología del Sistema NerviosoDe EverandAnatomía y Fisiología del Sistema NerviosoCalificación: 4 de 5 estrellas4/5 (19)
- Funcion Motora MedularDocumento53 páginasFuncion Motora MedularAlejandro D. Pérez GuillénAún no hay calificaciones
- Resumen Capitulo 55Documento8 páginasResumen Capitulo 55Rashel MoscosoAún no hay calificaciones
- Cap 54, Funciones Motoras de La Médula EspinalDocumento7 páginasCap 54, Funciones Motoras de La Médula EspinalPablo RosaAún no hay calificaciones
- De La Médula, y Otra Rama Transmite Sus Impulsos Hacia Niveles Más Altos Del Sistema Nervioso, EsDocumento7 páginasDe La Médula, y Otra Rama Transmite Sus Impulsos Hacia Niveles Más Altos Del Sistema Nervioso, EsJhony PradelaAún no hay calificaciones
- Reflejos FisiologiaDocumento24 páginasReflejos FisiologiaVictor MontesAún no hay calificaciones
- Funciones Motoras de La Médula EspinalDocumento12 páginasFunciones Motoras de La Médula EspinalBetsy VacaAún no hay calificaciones
- Clase 3 Médula Espinal, Reflejos y TonoDocumento25 páginasClase 3 Médula Espinal, Reflejos y Tonokhendra matusAún no hay calificaciones
- Médula EspinalDocumento94 páginasMédula EspinalJosé Nataniel Benítez SegoviaAún no hay calificaciones
- Capítulo 54 - GuytonDocumento4 páginasCapítulo 54 - GuytonJesica AndanAún no hay calificaciones
- Fisiología Sistema Nervioso y Shock Espinal 1Documento11 páginasFisiología Sistema Nervioso y Shock Espinal 1Azcem NavarreteAún no hay calificaciones
- Funciones Motoras de La Medula EspinalDocumento12 páginasFunciones Motoras de La Medula EspinalfernandoAún no hay calificaciones
- Reflejos medulares y control motorDocumento43 páginasReflejos medulares y control motorSergio Paz100% (1)
- Cap. 55 Funciones Motoras de La Médula Espinal - Reflejos MedularesDocumento10 páginasCap. 55 Funciones Motoras de La Médula Espinal - Reflejos MedularesLinda Andreita Rosales Rosales0% (1)
- 55-Funciones Motoras de La Medula EspinalDocumento8 páginas55-Funciones Motoras de La Medula EspinaledinAún no hay calificaciones
- 55-Funciones Motoras de La Medula EspinalDocumento8 páginas55-Funciones Motoras de La Medula EspinaledinAún no hay calificaciones
- Plantilla - Apunte Morado - StudyWithArtDocumento9 páginasPlantilla - Apunte Morado - StudyWithArtDENISE ALEJANDRA MARTINEZ GARCIAAún no hay calificaciones
- Reflejos EspinalesDocumento25 páginasReflejos EspinalesSchneiderAlvarezAún no hay calificaciones
- Capitulo 54 Resumen FisioDocumento6 páginasCapitulo 54 Resumen FisioFreddy TorresAún no hay calificaciones
- Capitulo 55Documento5 páginasCapitulo 55leslie laraAún no hay calificaciones
- Apuntes de FisiologíaDocumento8 páginasApuntes de FisiologíaOmar C. RodríguezAún no hay calificaciones
- 5 - Funciones Motoras de La MédulaDocumento53 páginas5 - Funciones Motoras de La MédulaDenner AfonsoAún no hay calificaciones
- Tema 19. Regulación MuscularDocumento21 páginasTema 19. Regulación MuscularYasmin BustosAún no hay calificaciones
- Funciones motoras de la médula espinal: reflejos medularesDocumento10 páginasFunciones motoras de la médula espinal: reflejos medularesEliezer Sop0% (1)
- M.E Funciones MotDocumento11 páginasM.E Funciones MotgeralAún no hay calificaciones
- OtgDocumento32 páginasOtgHugo Esteban LopezAún no hay calificaciones
- Preguntas Capitulo 54Documento12 páginasPreguntas Capitulo 54RobertoAndresGomezAún no hay calificaciones
- Capitulo 54 Funciones Motoras de La Medula EspinalDocumento10 páginasCapitulo 54 Funciones Motoras de La Medula EspinalMiguel MoraAún no hay calificaciones
- Neuro GeneralidadesDocumento5 páginasNeuro GeneralidadesEvangelina OviedoAún no hay calificaciones
- Reflejos MedularesDocumento5 páginasReflejos MedularesClinton anderson Luque mamaniAún no hay calificaciones
- Funciones Motoras de La Medula Espinal.Documento4 páginasFunciones Motoras de La Medula Espinal.Britney Aislinn VictorianoAún no hay calificaciones
- Funciones Motoras de La Médula EspinalDocumento23 páginasFunciones Motoras de La Médula EspinalLiss PeñafielAún no hay calificaciones
- Funciones Motoras de La Médula EspinalDocumento74 páginasFunciones Motoras de La Médula EspinalLourdes CalijauAún no hay calificaciones
- Médula EspinalDocumento83 páginasMédula EspinalNORA GUADALUPE SANCHEZ LORENZOAún no hay calificaciones
- FisiologiaDocumento22 páginasFisiologiaRafaela VeigaAún no hay calificaciones
- Fisio II Capítulo 55PR23Documento40 páginasFisio II Capítulo 55PR23BRYAN ALEJANDRO GONZALES CAMPOSAún no hay calificaciones
- Actividad Prática Nº5 - SapoDocumento12 páginasActividad Prática Nº5 - SapoDaniela AlejandraAún no hay calificaciones
- Fisiología - Reflejos MedularesDocumento7 páginasFisiología - Reflejos MedularesguafdalboraAún no hay calificaciones
- Fisiologia ReflejosDocumento6 páginasFisiologia ReflejosAGUSTÍN LEANDRO DÍAZAún no hay calificaciones
- Neurofisiologia Del MovimientoDocumento21 páginasNeurofisiologia Del MovimientoAlex Lopez0% (1)
- Morfo NerviosoDocumento22 páginasMorfo Nerviosocochja.19082003Aún no hay calificaciones
- Funciones motoras de la médula espinal: los reflejos medularesDocumento24 páginasFunciones motoras de la médula espinal: los reflejos medularesPaola AAún no hay calificaciones
- Síndrome de Motoneurona Superior e Inferior Final 2012Documento14 páginasSíndrome de Motoneurona Superior e Inferior Final 2012Diana FonsecaAún no hay calificaciones
- Control Del MovimientoDocumento28 páginasControl Del MovimientoMary Nicole Aráuz AráuzAún no hay calificaciones
- Enfermedades MotoneuronasDocumento94 páginasEnfermedades Motoneuronaskbautista20210215Aún no hay calificaciones
- 3.1neurofisiologiamotoraeintegradora 131009094438 Phpapp02 (1) Aec794627e668a7599c7e0193dc56710Documento20 páginas3.1neurofisiologiamotoraeintegradora 131009094438 Phpapp02 (1) Aec794627e668a7599c7e0193dc56710carlosAún no hay calificaciones
- Funciones Motoras de La Medula Espinal y ReflejosDocumento62 páginasFunciones Motoras de La Medula Espinal y ReflejosJoaoFranciscoEuphrasio100% (2)
- Cap 55 GuytonDocumento4 páginasCap 55 GuytonD-038 Ledis Melissa Quiñonez AranaAún no hay calificaciones
- Terminaciones Receptoras2Documento4 páginasTerminaciones Receptoras2Mavi MejiaAún no hay calificaciones
- Huso Muscular y OtgDocumento2 páginasHuso Muscular y OtgFelipe Mariano Ramos RuzAún no hay calificaciones
- Notas de Clases de NeuropsicologíaDocumento46 páginasNotas de Clases de NeuropsicologíamarielaAún no hay calificaciones
- Funciones Motoras de La Medula EspinalDocumento31 páginasFunciones Motoras de La Medula EspinalKarito Escobar P100% (1)
- Huso NeuromuscularDocumento33 páginasHuso NeuromuscularGab GahanAún no hay calificaciones
- Sistema MotorDocumento5 páginasSistema MotorMariana VargasAún no hay calificaciones
- Vías Motoras Voluntarias y Núcleos de La Base CompletitoDocumento20 páginasVías Motoras Voluntarias y Núcleos de La Base CompletitoAlvaro Alarcon ValenzuelaAún no hay calificaciones
- Captura de Pantalla 2023-11-11 A La(s) 21.43.55Documento16 páginasCaptura de Pantalla 2023-11-11 A La(s) 21.43.55MarcelaAún no hay calificaciones
- Capítulo 46Documento6 páginasCapítulo 46Rashel MoscosoAún no hay calificaciones
- Tarea Unidad 8 FISIOLOGIA II 2021 Funciones Motoras de La Medula EspinalDocumento3 páginasTarea Unidad 8 FISIOLOGIA II 2021 Funciones Motoras de La Medula EspinalNayeli PeñaAún no hay calificaciones
- Funciones de Los Ganglios BasalesDocumento13 páginasFunciones de Los Ganglios BasalesErwin RodriguezAún no hay calificaciones
- Calcio y FosfatoDocumento24 páginasCalcio y FosfatoErwin RodriguezAún no hay calificaciones
- Vitamina D, PTH y calcitoninaDocumento21 páginasVitamina D, PTH y calcitoninaErwin RodriguezAún no hay calificaciones
- Facultad de Medicina UA - Sistemas sensitivosDocumento20 páginasFacultad de Medicina UA - Sistemas sensitivosErwin RodriguezAún no hay calificaciones
- Demencia y EsquizofreniaDocumento18 páginasDemencia y EsquizofreniaErwin RodriguezAún no hay calificaciones
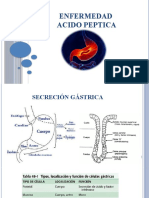
- Acido Peptica 2Documento48 páginasAcido Peptica 2Erwin RodriguezAún no hay calificaciones
- 2 Dinámica de La Membrana Celular 2015Documento47 páginas2 Dinámica de La Membrana Celular 2015Erwin RodriguezAún no hay calificaciones
- Los Sentidos QuímicosDocumento34 páginasLos Sentidos QuímicosErwin RodriguezAún no hay calificaciones
- Hia ScaDocumento37 páginasHia ScaErwin RodriguezAún no hay calificaciones
- Patologia Quirurgica Del HigadoDocumento45 páginasPatologia Quirurgica Del HigadoErwin RodriguezAún no hay calificaciones
- Hemostasia: Formación de Tapón PlaquetarioDocumento27 páginasHemostasia: Formación de Tapón PlaquetarioErwin RodriguezAún no hay calificaciones
- CONFERENCIA Isquemia Arterial AgudaDocumento42 páginasCONFERENCIA Isquemia Arterial AgudaErwin RodriguezAún no hay calificaciones
- Conferencia VaricesDocumento51 páginasConferencia VaricesErwin RodriguezAún no hay calificaciones
- AA Perforaciones DuodenalesDocumento21 páginasAA Perforaciones DuodenalesErwin RodriguezAún no hay calificaciones
- Revista - Introduccion A Neurociencias - 2022Documento9 páginasRevista - Introduccion A Neurociencias - 2022JheremyQenmAún no hay calificaciones
- TAC de cráneo y angiotomografía de cuello sin hallazgos patológicosDocumento2 páginasTAC de cráneo y angiotomografía de cuello sin hallazgos patológicosJaime Hernando Camacho pinzonAún no hay calificaciones
- Brainbox - f2 - Mod 6 - Tema 2Documento6 páginasBrainbox - f2 - Mod 6 - Tema 2Alejandra MoraAún no hay calificaciones
- Neuropa Tia Del Nervi o Puden DoDocumento11 páginasNeuropa Tia Del Nervi o Puden DoCristian Felipe Correa GallegoAún no hay calificaciones
- Actividad 2 - Atención y Procesos Psicológicos Superiores TODASDocumento28 páginasActividad 2 - Atención y Procesos Psicológicos Superiores TODASMARÍA ALEJANDRA DÍAZ MUÑOZAún no hay calificaciones
- Fisiología de La MicciónDocumento18 páginasFisiología de La MicciónDaniel RamírezAún no hay calificaciones
- Funciones Del Tronco EncefalicoDocumento4 páginasFunciones Del Tronco EncefalicoJesus MaresAún no hay calificaciones
- Guia Repaso Anatomía Cpu MarzoDocumento19 páginasGuia Repaso Anatomía Cpu MarzoMeei Osorio LimoAún no hay calificaciones
- Punteo de ClasesDocumento20 páginasPunteo de ClasesCarla RiveroAún no hay calificaciones
- Estrategias y Abordajes en Neurocirugía CranealDocumento610 páginasEstrategias y Abordajes en Neurocirugía CranealAyi Bastias100% (1)
- Manual Pato II-2021Documento143 páginasManual Pato II-2021David ArteagaAún no hay calificaciones
- ACTIVACION de La GLANDULA PINEALDocumento4 páginasACTIVACION de La GLANDULA PINEALliliana.amielli100% (1)
- Partes Del Cerebro y Su FunciónDocumento2 páginasPartes Del Cerebro y Su FunciónalejandraAún no hay calificaciones
- Glosario SemiologiaDocumento18 páginasGlosario SemiologiaErika CuencaAún no hay calificaciones
- SindromeDocumento3 páginasSindromeYANIRE NATALIE SIFUENTES ZUÑIGAAún no hay calificaciones
- Actividad #6Documento7 páginasActividad #6Florencia BillordoAún no hay calificaciones
- Guía de Estudio 4. Homeostasis, Medio Interno, Ph. Planimetría, Regiones y Cavidades Del CuerpoDocumento17 páginasGuía de Estudio 4. Homeostasis, Medio Interno, Ph. Planimetría, Regiones y Cavidades Del CuerpoGloriaAún no hay calificaciones
- Resumen Segundo Examen de Fisiologia Animal, Guillermo CastroDocumento39 páginasResumen Segundo Examen de Fisiologia Animal, Guillermo CastroRAMON FLORESAún no hay calificaciones
- Temario AdultoDocumento82 páginasTemario AdultoVerónica GetteAún no hay calificaciones
- Tejido Nervioso (Ganglios Nerviosos) - Alfredo Rubiano CaballeroDocumento6 páginasTejido Nervioso (Ganglios Nerviosos) - Alfredo Rubiano CaballeroCarolina RodriguezAún no hay calificaciones
- Simulacion de Linfomas y Aparato GastrointestinalDocumento132 páginasSimulacion de Linfomas y Aparato GastrointestinalNancy Pedroza MendezAún no hay calificaciones
- Lista de Rastreo SoftwareDocumento26 páginasLista de Rastreo SoftwareRolando Bravo100% (76)
- Embriología Del Tubo Digestivo (Clase Gastro)Documento57 páginasEmbriología Del Tubo Digestivo (Clase Gastro)elizabeth lmAún no hay calificaciones
- ACTIVIDAD #20 - 3°-Ra - SEM.-24-CISNEROSDocumento5 páginasACTIVIDAD #20 - 3°-Ra - SEM.-24-CISNEROSyosiAún no hay calificaciones
- Pae Aneurisma FinalDocumento36 páginasPae Aneurisma FinalDeni RivSant100% (1)
- Origen y Desarrollo Del SNADocumento34 páginasOrigen y Desarrollo Del SNAXbrutalX2Aún no hay calificaciones
- Taller 5 Sistema NerviosoDocumento29 páginasTaller 5 Sistema NerviosoleidyAún no hay calificaciones
- Lección - EVALUACIÓN PRIMARIADocumento6 páginasLección - EVALUACIÓN PRIMARIAJason Alexander Zutara MoralesAún no hay calificaciones
- Nervio OpticoDocumento2 páginasNervio OpticoDiana RosasAún no hay calificaciones
- Grant Anatomia para ColorearDocumento559 páginasGrant Anatomia para ColorearLuiz Felipe Gonçalves100% (1)
- Neurociencia para vencer la depresión: La esprial ascendenteDe EverandNeurociencia para vencer la depresión: La esprial ascendenteCalificación: 4.5 de 5 estrellas4.5/5 (10)
- Cómo conversar con cualquiera: Descubre cómo ser la persona más interesante del lugar y a destruir la ansiedad social permanentementeDe EverandCómo conversar con cualquiera: Descubre cómo ser la persona más interesante del lugar y a destruir la ansiedad social permanentementeCalificación: 4 de 5 estrellas4/5 (81)
- Terapia cognitiva: Conceptos básicos y profundizaciónDe EverandTerapia cognitiva: Conceptos básicos y profundizaciónCalificación: 5 de 5 estrellas5/5 (1)
- Resumen de Pensar rápido pensar despacio de Daniel KahnemanDe EverandResumen de Pensar rápido pensar despacio de Daniel KahnemanCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Dinámicas familiares a través de la vida de José: La túnica del PadreDe EverandDinámicas familiares a través de la vida de José: La túnica del PadreCalificación: 5 de 5 estrellas5/5 (2)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Terapia cognitiva del comportamiento: Descubre cómo despejar tu cerebro con la TCC. Aprender a superar el miedo y la ansiedad, la depresiónDe EverandTerapia cognitiva del comportamiento: Descubre cómo despejar tu cerebro con la TCC. Aprender a superar el miedo y la ansiedad, la depresiónCalificación: 5 de 5 estrellas5/5 (3)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- Psicología y trastornos de los niños en la edad evolutiva: Qué son y cómo funcionanDe EverandPsicología y trastornos de los niños en la edad evolutiva: Qué son y cómo funcionanCalificación: 5 de 5 estrellas5/5 (1)
- DMT: La molécula del espíritu: Las revolucionarias investigaciones de un médico sobre la biología de las experiencias místicas y cercanas a la muerteDe EverandDMT: La molécula del espíritu: Las revolucionarias investigaciones de un médico sobre la biología de las experiencias místicas y cercanas a la muerteCalificación: 4 de 5 estrellas4/5 (135)
- Bioneuroemoción: Un método para el bienestar emocionalDe EverandBioneuroemoción: Un método para el bienestar emocionalCalificación: 5 de 5 estrellas5/5 (4)
- Cómo estimular el cerebro del niño: 100 ejercicios para potenciar la concentración, la memoria y otras funciones ejecutivasDe EverandCómo estimular el cerebro del niño: 100 ejercicios para potenciar la concentración, la memoria y otras funciones ejecutivasCalificación: 4.5 de 5 estrellas4.5/5 (12)
- Notas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesDe EverandNotas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesAún no hay calificaciones
- Mente Organizada Cómo Pensar con Claridad y Hacer Todas Las Decisiones de Vida CorrectasDe EverandMente Organizada Cómo Pensar con Claridad y Hacer Todas Las Decisiones de Vida CorrectasCalificación: 4 de 5 estrellas4/5 (11)
- El cerebro del niño explicado a los padresDe EverandEl cerebro del niño explicado a los padresCalificación: 4.5 de 5 estrellas4.5/5 (147)
- El arte del descanso: Descubre el método para dormir bien y descansar mejorDe EverandEl arte del descanso: Descubre el método para dormir bien y descansar mejorCalificación: 4.5 de 5 estrellas4.5/5 (4)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- 200 tareas en terapia breve: 2ª ediciónDe Everand200 tareas en terapia breve: 2ª ediciónCalificación: 4.5 de 5 estrellas4.5/5 (33)
- Mindfulness: Cómo hacer menos y ser más conscientes puede ayudarnos a vivir mejor en este mundo estresadoDe EverandMindfulness: Cómo hacer menos y ser más conscientes puede ayudarnos a vivir mejor en este mundo estresadoCalificación: 4.5 de 5 estrellas4.5/5 (9)
- Guía para superar los pensamientos atemorizantes, obsesivos o inquietantes: Libérate de los pensamientos negativos con la terapia cognitivo conductual (CBT)De EverandGuía para superar los pensamientos atemorizantes, obsesivos o inquietantes: Libérate de los pensamientos negativos con la terapia cognitivo conductual (CBT)Calificación: 4 de 5 estrellas4/5 (25)
- La conciencia en el cerebro: Descifrando el enigma de cómo el cerebro elabora nuestros pensamientosDe EverandLa conciencia en el cerebro: Descifrando el enigma de cómo el cerebro elabora nuestros pensamientosCalificación: 2.5 de 5 estrellas2.5/5 (3)
- Camino hacia el sí mismo. Las cuatro facetas humanas desde una psicología integrativa: Lo cognitivo, lo emocional, lo existencial, lo espiritualDe EverandCamino hacia el sí mismo. Las cuatro facetas humanas desde una psicología integrativa: Lo cognitivo, lo emocional, lo existencial, lo espiritualCalificación: 5 de 5 estrellas5/5 (1)
- Abrázame fuerte: Siete conversaciones para un amor duraderoDe EverandAbrázame fuerte: Siete conversaciones para un amor duraderoManu BerásteguiCalificación: 4.5 de 5 estrellas4.5/5 (13)
- Cerebros rotos: Pacientes asombrosos que me enseñaron a vivirDe EverandCerebros rotos: Pacientes asombrosos que me enseñaron a vivirCalificación: 5 de 5 estrellas5/5 (3)