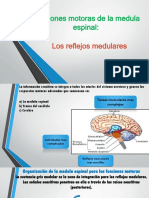
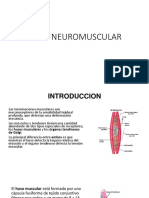
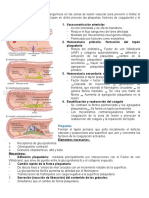
También podría gustarte
- Funciones Motoras de La Medula EspinalDocumento31 páginasFunciones Motoras de La Medula EspinalKarito Escobar P100% (1)
- Funciones Motoras de La Medula EspinalDocumento12 páginasFunciones Motoras de La Medula EspinalfernandoAún no hay calificaciones
- Anatomía y Fisiología del Sistema NerviosoDe EverandAnatomía y Fisiología del Sistema NerviosoCalificación: 4 de 5 estrellas4/5 (19)
- Pantalla de PilasDocumento58 páginasPantalla de PilasJOSÉ ENRIQUE ARENAS CORPASAún no hay calificaciones
- Cartografia ComunitariaDocumento59 páginasCartografia ComunitariaLeo BiberAún no hay calificaciones
- Perfil Centro CivicoDocumento92 páginasPerfil Centro CivicoShanyi Leandro BayonaAún no hay calificaciones
- Diagnostico Situacional CAPDocumento7 páginasDiagnostico Situacional CAPABy ArriazaAún no hay calificaciones
- Resumen Capitulo 55Documento8 páginasResumen Capitulo 55Rashel MoscosoAún no hay calificaciones
- Funciones Motoras de La Medula Espinal Reflejos Medulares DR MerelloDocumento83 páginasFunciones Motoras de La Medula Espinal Reflejos Medulares DR Merellojosemerello100% (1)
- Funcion Motora MedularDocumento53 páginasFuncion Motora MedularAlejandro D. Pérez GuillénAún no hay calificaciones
- Reflejos medulares y control motorDocumento43 páginasReflejos medulares y control motorSergio Paz100% (1)
- Funciones motoras de la médula espinal y reflejosDocumento77 páginasFunciones motoras de la médula espinal y reflejosErwin RodriguezAún no hay calificaciones
- Funciones Motoras de La Médula EspinalDocumento12 páginasFunciones Motoras de La Médula EspinalBetsy VacaAún no hay calificaciones
- Capitulo 55Documento5 páginasCapitulo 55leslie laraAún no hay calificaciones
- Cap 55. Función Motora de La Médula Espinal, Los Reflejos MedularesDocumento4 páginasCap 55. Función Motora de La Médula Espinal, Los Reflejos MedularesDaniela SolísAún no hay calificaciones
- Cap 54, Funciones Motoras de La Médula EspinalDocumento7 páginasCap 54, Funciones Motoras de La Médula EspinalPablo RosaAún no hay calificaciones
- De La Médula, y Otra Rama Transmite Sus Impulsos Hacia Niveles Más Altos Del Sistema Nervioso, EsDocumento7 páginasDe La Médula, y Otra Rama Transmite Sus Impulsos Hacia Niveles Más Altos Del Sistema Nervioso, EsJhony PradelaAún no hay calificaciones
- Fisiología - Reflejos MedularesDocumento7 páginasFisiología - Reflejos MedularesguafdalboraAún no hay calificaciones
- 55-Funciones Motoras de La Medula EspinalDocumento8 páginas55-Funciones Motoras de La Medula EspinaledinAún no hay calificaciones
- 55-Funciones Motoras de La Medula EspinalDocumento8 páginas55-Funciones Motoras de La Medula EspinaledinAún no hay calificaciones
- Reflejo Medular: ClasificaciónDocumento6 páginasReflejo Medular: ClasificaciónJoab Lira DiazAún no hay calificaciones
- Neuro GeneralidadesDocumento5 páginasNeuro GeneralidadesEvangelina OviedoAún no hay calificaciones
- Médula EspinalDocumento94 páginasMédula EspinalJosé Nataniel Benítez SegoviaAún no hay calificaciones
- Reflejos FisiologiaDocumento24 páginasReflejos FisiologiaVictor MontesAún no hay calificaciones
- 5 - Funciones Motoras de La MédulaDocumento53 páginas5 - Funciones Motoras de La MédulaDenner AfonsoAún no hay calificaciones
- NEUROFISIOLOGIA Funciones Mototras 2022Documento25 páginasNEUROFISIOLOGIA Funciones Mototras 2022Edgardo RamirezAún no hay calificaciones
- Clase 3 Médula Espinal, Reflejos y TonoDocumento25 páginasClase 3 Médula Espinal, Reflejos y Tonokhendra matusAún no hay calificaciones
- Plantilla - Apunte Morado - StudyWithArtDocumento9 páginasPlantilla - Apunte Morado - StudyWithArtDENISE ALEJANDRA MARTINEZ GARCIAAún no hay calificaciones
- 7 Organizacion de La Medula Espinal para Las Funciones MotorasDocumento52 páginas7 Organizacion de La Medula Espinal para Las Funciones MotorasKary Araujo100% (2)
- Funciones Motoras de La Medula Espinal y ReflejosDocumento62 páginasFunciones Motoras de La Medula Espinal y ReflejosJoaoFranciscoEuphrasio100% (2)
- Reflejo MedularDocumento5 páginasReflejo MedularMaily MuñozAún no hay calificaciones
- Tema 19. Regulación MuscularDocumento21 páginasTema 19. Regulación MuscularYasmin BustosAún no hay calificaciones
- Huso NeuromuscularDocumento29 páginasHuso NeuromuscularaaaaAún no hay calificaciones
- Funciones Motoras de La Médula EspinalDocumento74 páginasFunciones Motoras de La Médula EspinalLourdes CalijauAún no hay calificaciones
- Reflejos EspinalesDocumento25 páginasReflejos EspinalesSchneiderAlvarezAún no hay calificaciones
- Sensibilidad PropioceptivaDocumento11 páginasSensibilidad PropioceptivaKarla ArriagadaAún no hay calificaciones
- Capitulo 54 Resumen FisioDocumento6 páginasCapitulo 54 Resumen FisioFreddy TorresAún no hay calificaciones
- Sistema Inhibidor de La Celula de RenshawDocumento8 páginasSistema Inhibidor de La Celula de RenshawMonte Sann MarielaAún no hay calificaciones
- Médula EspinalDocumento83 páginasMédula EspinalNORA GUADALUPE SANCHEZ LORENZOAún no hay calificaciones
- Tonos musculares y sus implicaciones en el desarrollo motorDocumento33 páginasTonos musculares y sus implicaciones en el desarrollo motorRaúl Nolasco VelazquezAún no hay calificaciones
- Vías Motoras Voluntarias y Núcleos de La Base CompletitoDocumento20 páginasVías Motoras Voluntarias y Núcleos de La Base CompletitoAlvaro Alarcon ValenzuelaAún no hay calificaciones
- Apuntes de FisiologíaDocumento8 páginasApuntes de FisiologíaOmar C. RodríguezAún no hay calificaciones
- M.E Funciones MotDocumento11 páginasM.E Funciones MotgeralAún no hay calificaciones
- Organizacion Jerarquica Del SNCDocumento69 páginasOrganizacion Jerarquica Del SNCPamela Mejia de Melo75% (4)
- Reflejo miotático: el reflejo de estiramiento muscularDocumento8 páginasReflejo miotático: el reflejo de estiramiento muscularMaria RamosAún no hay calificaciones
- Organizac. SN. Reflejos Medulares1Documento41 páginasOrganizac. SN. Reflejos Medulares1Yenisei Faustinos GutierrezAún no hay calificaciones
- Reflejos MedularesDocumento5 páginasReflejos MedularesClinton anderson Luque mamaniAún no hay calificaciones
- Función Motora de La Medula Espinal - Reflejos MedularesDocumento21 páginasFunción Motora de La Medula Espinal - Reflejos MedularesAlejandro Montiel CortezAún no hay calificaciones
- Funciones motoras de la médula espinal: reflejos medularesDocumento10 páginasFunciones motoras de la médula espinal: reflejos medularesEliezer Sop0% (1)
- 21 FisioDocumento8 páginas21 FisioDaniela Kohbra Castro MabhootAún no hay calificaciones
- Reflejo MedularDocumento6 páginasReflejo MedularNatalie MitreAún no hay calificaciones
- Actividad Prática Nº5 - SapoDocumento12 páginasActividad Prática Nº5 - SapoDaniela AlejandraAún no hay calificaciones
- Reflejos espinales y posturales: funciones y mecanismosDocumento29 páginasReflejos espinales y posturales: funciones y mecanismosMIGUEL ANGEL PALOMINO CAMPOAún no hay calificaciones
- Capitulo 54 Funciones Motoras de La Medula EspinalDocumento10 páginasCapitulo 54 Funciones Motoras de La Medula EspinalMiguel MoraAún no hay calificaciones
- Tono muscular: regulación y componentes del reflejo miotáticoDocumento10 páginasTono muscular: regulación y componentes del reflejo miotáticoDaniel Aaron Fernandez GuzmanAún no hay calificaciones
- OtgDocumento32 páginasOtgHugo Esteban LopezAún no hay calificaciones
- Clase 2.medula EspinalDocumento41 páginasClase 2.medula EspinalJosebeth RisquezAún no hay calificaciones
- Cap. 55 Funciones Motoras de La Médula Espinal - Reflejos MedularesDocumento10 páginasCap. 55 Funciones Motoras de La Médula Espinal - Reflejos MedularesLinda Andreita Rosales Rosales0% (1)
- Mecanismo NeuromuscularDocumento10 páginasMecanismo NeuromuscularWenMuñoz100% (1)
- Clase 2-FISIODocumento10 páginasClase 2-FISIOmilenka.olivares.sanhuezaAún no hay calificaciones
- Sensibilidad PropioceptivaDocumento22 páginasSensibilidad PropioceptivaMichael DuranAún no hay calificaciones
- 3.2.sensibilidad PropioceptivaDocumento12 páginas3.2.sensibilidad PropioceptivaDony ZamoranoAún no hay calificaciones
- Fisio II Capítulo 55PR23Documento40 páginasFisio II Capítulo 55PR23BRYAN ALEJANDRO GONZALES CAMPOSAún no hay calificaciones
- Anatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2De EverandAnatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2Aún no hay calificaciones
- Entrevista Hombrecito de Defensa CivilDocumento1 páginaEntrevista Hombrecito de Defensa CivilGrecia M CoronadoAún no hay calificaciones
- Homunculos CorticalesDocumento2 páginasHomunculos CorticalesVanesa Camila Málaga SalcedoAún no hay calificaciones
- Introduccion A La ComunicaciónDocumento12 páginasIntroduccion A La ComunicaciónFreddy Cruz Segovia100% (1)
- Cultura ChavinDocumento2 páginasCultura ChavinSebastian GallegosAún no hay calificaciones
- Fuerza Radial o Cent Rip EtaDocumento5 páginasFuerza Radial o Cent Rip EtakmantillaAún no hay calificaciones
- Actividades CNBDocumento32 páginasActividades CNBClaudita NolascoAún no hay calificaciones
- Proyec Invest QarqanchaDocumento8 páginasProyec Invest QarqanchaangeltoralvaAún no hay calificaciones
- Ciencias Naturales Plan Formativo 5to PrimariaDocumento7 páginasCiencias Naturales Plan Formativo 5to PrimariaCarlos PinedaAún no hay calificaciones
- HidrogramaDocumento21 páginasHidrogramaJerson Cuba Sanchez100% (1)
- Anatomía de las nalgasDocumento1 páginaAnatomía de las nalgasDalina Cortez DTAún no hay calificaciones
- Georges Louis Leclerc, naturalista francés (1707-1788Documento7 páginasGeorges Louis Leclerc, naturalista francés (1707-1788Frodo NenAún no hay calificaciones
- Miralles Y SUS COSAS PDFDocumento247 páginasMiralles Y SUS COSAS PDFCarlos SAnchez Raggini100% (2)
- Prueba Historia Segundo Basico - Ensayos de Calidad - Smsg0325Documento2 páginasPrueba Historia Segundo Basico - Ensayos de Calidad - Smsg0325rousesAún no hay calificaciones
- La Tierra en Clave de SistemaDocumento1 páginaLa Tierra en Clave de SistemaNathalia two thousand and oneAún no hay calificaciones
- Serranía EstepariaDocumento7 páginasSerranía EstepariaAdriana Carranza0% (1)
- Homeostasis coagulaciónDocumento3 páginasHomeostasis coagulaciónYesenia JimenezAún no hay calificaciones
- Evaluacion Ambiental Estrategica Del Corredor Norte de Bolivia DiagnosticoDocumento235 páginasEvaluacion Ambiental Estrategica Del Corredor Norte de Bolivia DiagnosticoBernie_Garcia__9886Aún no hay calificaciones
- MONOGRAFIADocumento43 páginasMONOGRAFIAsassyta90c100% (2)
- Técnicas de Contaje Celular NeubauerDocumento2 páginasTécnicas de Contaje Celular NeubauerResrod78Aún no hay calificaciones
- Laboratorio Nº1 (Grasas y Aceite)Documento4 páginasLaboratorio Nº1 (Grasas y Aceite)JoseMiguelVediaRochaAún no hay calificaciones
- El Discurso Steve Jobs, EmprendimientoDocumento7 páginasEl Discurso Steve Jobs, Emprendimientocramirezrojas@yahoo.comAún no hay calificaciones
- Sergio Nicolas Salas 2015-101039Documento10 páginasSergio Nicolas Salas 2015-101039Rut ZapanaAún no hay calificaciones
- Conductismo: Estudio de la conducta observableDocumento10 páginasConductismo: Estudio de la conducta observableFranco Velazco VentoAún no hay calificaciones
- Ensayo de Placa EstáticoDocumento9 páginasEnsayo de Placa EstáticoFredy OrtizAún no hay calificaciones
- Como Se Formaron Los PlanetasDocumento2 páginasComo Se Formaron Los PlanetasAlejandra LopezAún no hay calificaciones
- Los Hongos Son Seres HeterótrofosDocumento33 páginasLos Hongos Son Seres HeterótrofosLinford Kits Porta ChAún no hay calificaciones