También podría gustarte
- Introducción a la Biología: RESÚMENES UNIVERSITARIOSDe EverandIntroducción a la Biología: RESÚMENES UNIVERSITARIOSCalificación: 5 de 5 estrellas5/5 (1)
- Cuestionario LCTDocumento17 páginasCuestionario LCTMichel Gonzalez100% (1)
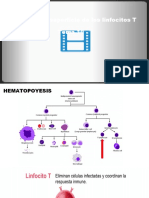
- Desarrollo TCR linfocitos TDocumento9 páginasDesarrollo TCR linfocitos TRubén HAún no hay calificaciones
- Clase 7 - Ontogenia de Linfocitos TDocumento10 páginasClase 7 - Ontogenia de Linfocitos TLautaro VasqueAún no hay calificaciones
- TCR 1Documento14 páginasTCR 1Kevin alberto Pérez de la cruzAún no hay calificaciones
- Desarrollo de Linfocitos TDocumento2 páginasDesarrollo de Linfocitos Ta358191Aún no hay calificaciones
- clase-4-linfocitos-t-2Documento13 páginasclase-4-linfocitos-t-2fialophora0210Aún no hay calificaciones
- 07 - Ontogenia TDocumento30 páginas07 - Ontogenia T25984076Aún no hay calificaciones
- Moléculas de Superficie de Los LTDocumento44 páginasMoléculas de Superficie de Los LTLeninReyesAún no hay calificaciones
- Linfocitos T y receptores de membranaDocumento20 páginasLinfocitos T y receptores de membranaMELISSA JOHANA GONZALEZ SALCEDOAún no hay calificaciones
- Ontogenia de Los Linfocitos TDocumento6 páginasOntogenia de Los Linfocitos TYamiiLette CorniizAún no hay calificaciones
- LinfocitosDocumento1 páginaLinfocitosPatricia NeriAún no hay calificaciones
- Ontogenia T y selección linfocitos TDocumento25 páginasOntogenia T y selección linfocitos TMaría Melina CedrúnAún no hay calificaciones
- BCR y TCRDocumento12 páginasBCR y TCRPaúl HaroAún no hay calificaciones
- Linfocitos T y Su Receptor (TCR)Documento8 páginasLinfocitos T y Su Receptor (TCR)Giovanna B100% (11)
- Capítulo 13 Pavón. Linfocitos TDocumento12 páginasCapítulo 13 Pavón. Linfocitos TClaudia PérezAún no hay calificaciones
- TEMA 10 Sin AudioDocumento25 páginasTEMA 10 Sin AudioNorelys SantosAún no hay calificaciones
- Apunte 03-11 Maduración Células TDocumento3 páginasApunte 03-11 Maduración Células TMichelle Alejandra Rivera GarciaAún no hay calificaciones
- 04 Linfocitos TDocumento37 páginas04 Linfocitos TNelson Quispe MaraniAún no hay calificaciones
- Linfocitos T y Su Receptor de Antígeno (TCR)Documento8 páginasLinfocitos T y Su Receptor de Antígeno (TCR)Mariano Félix DemasiAún no hay calificaciones
- Receptores de las células T: estructura y función del TCRDocumento19 páginasReceptores de las células T: estructura y función del TCRBrenda GonzalesAún no hay calificaciones
- Linfocitos TDocumento4 páginasLinfocitos TLucio WekslerAún no hay calificaciones
- Resumen Defensa-UP1 PDFDocumento57 páginasResumen Defensa-UP1 PDFIvan ChavesAún no hay calificaciones
- Linfocitos TDocumento15 páginasLinfocitos TIGNACIO SALVADOR FERNANDEZ GIRONAún no hay calificaciones
- Maduración de Linfocitos TDocumento41 páginasMaduración de Linfocitos TVicToRia50% (2)
- LINFOPOYESIS11Documento40 páginasLINFOPOYESIS11María Belén Silva VelásquezAún no hay calificaciones
- 2.1 Clases de HomeroDocumento34 páginas2.1 Clases de HomeroAndres Ignacio Aguilar AndradeAún no hay calificaciones
- Estructura y función del receptor de célula B y TCRDocumento9 páginasEstructura y función del receptor de célula B y TCRAnny ZhengAún no hay calificaciones
- Activación, diferenciación y memoria de células T: Receptores coestimuladoresDocumento14 páginasActivación, diferenciación y memoria de células T: Receptores coestimuladoresFabian Acosta ReyesAún no hay calificaciones
- Ontogenia Linfocitos TDocumento5 páginasOntogenia Linfocitos TmarianaaguillonAún no hay calificaciones
- Activacion de Linfocitos TDocumento18 páginasActivacion de Linfocitos TVictor Fabian Uribe Olmos100% (1)
- INMUNIDAD CELULAR ResumenDocumento16 páginasINMUNIDAD CELULAR ResumenwendyAún no hay calificaciones
- Maduración y Activación de Linfocitos TDocumento59 páginasMaduración y Activación de Linfocitos TVioleta Tello MedinaAún no hay calificaciones
- Preguntas para La Unidad 3 de Inmunología Referente A Órganos y Células LinfoidesDocumento4 páginasPreguntas para La Unidad 3 de Inmunología Referente A Órganos y Células LinfoidesSerCL1Aún no hay calificaciones
- INMUNIDAD CELULAR Estudiantes PDFDocumento49 páginasINMUNIDAD CELULAR Estudiantes PDFlordgimli77100% (3)
- Moléculas Presentadoras de AntígenoDocumento11 páginasMoléculas Presentadoras de AntígenoMariano Félix DemasiAún no hay calificaciones
- Tema 7 - Caracter¡sticas Generales de La L¡nea LinfocitariaDocumento20 páginasTema 7 - Caracter¡sticas Generales de La L¡nea LinfocitariaPablo GarciaAún no hay calificaciones
- Anticuerpos MonoclonicosDocumento10 páginasAnticuerpos MonoclonicosGabriela AlvarezAún no hay calificaciones
- Estructura: Co-Receptores T CD4 y CD8, Sinapsis Inmunológica. Receptores BCR y Co-Receptores Del Linfocito B - Grupo 7Documento7 páginasEstructura: Co-Receptores T CD4 y CD8, Sinapsis Inmunológica. Receptores BCR y Co-Receptores Del Linfocito B - Grupo 7Joseph Bryan Vera RodriguezAún no hay calificaciones
- Receptor Del LB y LT y OntogeniaDocumento5 páginasReceptor Del LB y LT y Ontogeniavalentinaxson2Aún no hay calificaciones
- Antígenos de Diferenciación LeucocitariaDocumento4 páginasAntígenos de Diferenciación LeucocitariaArmando LlvAún no hay calificaciones
- Ontogenia TDocumento5 páginasOntogenia TGiudiche BrisaAún no hay calificaciones
- Desarrollo de Células T en El TimoDocumento24 páginasDesarrollo de Células T en El TimoJames SantiagoAún no hay calificaciones
- Inmunidad TDocumento4 páginasInmunidad TCATHERINE MARÍA DE LOS ANGELES MAMANI APAZAAún no hay calificaciones
- 02 (Linfocitos T)Documento19 páginas02 (Linfocitos T)Francisco LedezmaAún no hay calificaciones
- Estructura y función del TCR y BCR en linfocitos T y BDocumento6 páginasEstructura y función del TCR y BCR en linfocitos T y BEsther MoralesAún no hay calificaciones
- Clase 3 Linfocito B y T SEMANA 3Documento64 páginasClase 3 Linfocito B y T SEMANA 3OCTAVIOAún no hay calificaciones
- Cap 10Documento7 páginasCap 10Natalia Espíndola BalderasAún no hay calificaciones
- Resumen Segundo Parcial (Marilin)Documento21 páginasResumen Segundo Parcial (Marilin)Nico Alister CortésAún no hay calificaciones
- Linfocitos TDocumento4 páginasLinfocitos TPaulaAlvarezSuaAún no hay calificaciones
- El complejo receptor TCR-CD3 y su papel en el reconocimiento antigénicoDocumento31 páginasEl complejo receptor TCR-CD3 y su papel en el reconocimiento antigénicoRosa PeñaAún no hay calificaciones
- CD markers for cell differentiationDocumento4 páginasCD markers for cell differentiationAngie Paola Sandoval CAún no hay calificaciones
- Linf T 1Documento69 páginasLinf T 1Alex David Sigueñas NoriegaAún no hay calificaciones
- Unidad 5 Fase de Reconocimiento, Activación y EfectoraDocumento29 páginasUnidad 5 Fase de Reconocimiento, Activación y EfectoralitorAún no hay calificaciones
- Clase 10. ONTOGENIA DE LAS CÉLULAS LINFOIDESDocumento6 páginasClase 10. ONTOGENIA DE LAS CÉLULAS LINFOIDESKevin VelaAún no hay calificaciones
- Telomeros y TelomerasasDocumento4 páginasTelomeros y TelomerasasLuz Vanessa ParejoAún no hay calificaciones
- Linfocitos T CD4+Documento12 páginasLinfocitos T CD4+pitu anatoAún no hay calificaciones
- Taller Linfopoyesis T y TCR Subgrupo 16Documento26 páginasTaller Linfopoyesis T y TCR Subgrupo 16RanciovichAún no hay calificaciones
- Clase 6 R. Linfocitos B y TDocumento25 páginasClase 6 R. Linfocitos B y TRene AlbertoAún no hay calificaciones
- Alimentos azucaradosDocumento3 páginasAlimentos azucaradosBioqui En ProcesoAún no hay calificaciones
- LB activaciónDocumento10 páginasLB activaciónBioqui En ProcesoAún no hay calificaciones
- Respuesta frente a helmintosDocumento2 páginasRespuesta frente a helmintosBioqui En ProcesoAún no hay calificaciones
- Reacciones de hipersensibilidadDocumento14 páginasReacciones de hipersensibilidadBioqui En ProcesoAún no hay calificaciones
- Mecanismos efectoresDocumento5 páginasMecanismos efectoresBioqui En ProcesoAún no hay calificaciones
- Actividad AcuosaDocumento7 páginasActividad AcuosaBioqui En ProcesoAún no hay calificaciones
- Dietas-conceptos generalesDocumento4 páginasDietas-conceptos generalesBioqui En ProcesoAún no hay calificaciones
- Infecciones Virales Genitourinarias: Virus Papiloma HumanoDocumento3 páginasInfecciones Virales Genitourinarias: Virus Papiloma HumanoBioqui En ProcesoAún no hay calificaciones
- 1-Fosfatasa LecturaDocumento1 página1-Fosfatasa LecturaBioqui En ProcesoAún no hay calificaciones
- APLICACIONESDocumento4 páginasAPLICACIONESBioqui En ProcesoAún no hay calificaciones
- 1-Fosfatasa ResultadosDocumento1 página1-Fosfatasa ResultadosBioqui En ProcesoAún no hay calificaciones
- Leyes de La RadioactividadDocumento2 páginasLeyes de La RadioactividadBioqui En ProcesoAún no hay calificaciones
- Espectrometria de MasasDocumento10 páginasEspectrometria de MasasBioqui En ProcesoAún no hay calificaciones
- Sistemas Automaticos y AutomatizadosDocumento2 páginasSistemas Automaticos y AutomatizadosBioqui En ProcesoAún no hay calificaciones
- TÉCNICAS VOLTAMPEROMÉTRICAS HIDRODINÁMICASDocumento3 páginasTÉCNICAS VOLTAMPEROMÉTRICAS HIDRODINÁMICASBioqui En ProcesoAún no hay calificaciones
- Efectos Químicos y BiológicosDocumento5 páginasEfectos Químicos y BiológicosBioqui En ProcesoAún no hay calificaciones
- Interacción Con La MateriaDocumento4 páginasInteracción Con La MateriaBioqui En ProcesoAún no hay calificaciones
- TÉCNICAS VOLTAMPEROMÉTRICAS HIDRODINÁMICASDocumento3 páginasTÉCNICAS VOLTAMPEROMÉTRICAS HIDRODINÁMICASBioqui En ProcesoAún no hay calificaciones
- Sistemas Automaticos y AutomatizadosDocumento2 páginasSistemas Automaticos y AutomatizadosBioqui En ProcesoAún no hay calificaciones
- Efectos Químicos y BiológicosDocumento5 páginasEfectos Químicos y BiológicosBioqui En ProcesoAún no hay calificaciones
- Interacción Con La MateriaDocumento4 páginasInteracción Con La MateriaBioqui En ProcesoAún no hay calificaciones
- Leyes de La RadioactividadDocumento2 páginasLeyes de La RadioactividadBioqui En ProcesoAún no hay calificaciones
- Espectrometria de MasasDocumento10 páginasEspectrometria de MasasBioqui En ProcesoAún no hay calificaciones
- Expo Donacion de OrganosDocumento15 páginasExpo Donacion de OrganosAnali Huanca HanccoAún no hay calificaciones
- KinesiologiaDocumento10 páginasKinesiologiaLuis RodriguezAún no hay calificaciones
- Que Es GanaderíaDocumento3 páginasQue Es GanaderíaWindar656Aún no hay calificaciones
- Identificación de Los Riesgos Aspecto QuimicoDocumento41 páginasIdentificación de Los Riesgos Aspecto QuimicoAs Mile Soler0% (1)
- Clasificacion BacterianaDocumento8 páginasClasificacion BacterianaCompartiendo Conocimientos MédicosAún no hay calificaciones
- Choque Septico Respuesta InmunologicaDocumento21 páginasChoque Septico Respuesta InmunologicaCristian André Mejía SalasAún no hay calificaciones
- Ventilacion MecanicaDocumento25 páginasVentilacion Mecanicaarpe100% (10)
- Cuentos Hermanos GrimmDocumento282 páginasCuentos Hermanos GrimmFredy Reinaldo Rivera MartínezAún no hay calificaciones
- Fisiologia Respiratoria Del Recien NacidoDocumento43 páginasFisiologia Respiratoria Del Recien NacidoAnonymous moiUr6100% (3)
- Tejido CartilaginosoDocumento3 páginasTejido CartilaginosoJosue Garrido MolinaAún no hay calificaciones
- Isquemia IntestinalDocumento10 páginasIsquemia IntestinalKoraima Villar CorzoAún no hay calificaciones
- Virus de La Hepatitis CDocumento6 páginasVirus de La Hepatitis CFrancis Vallecillo PadillaAún no hay calificaciones
- Alimentos Nutritivo2Documento5 páginasAlimentos Nutritivo2Marina Hua CenAún no hay calificaciones
- Animales de Compania PDFDocumento16 páginasAnimales de Compania PDFMonica Rodriguez CalderonAún no hay calificaciones
- Leccion 2. EMBRIOLOGIA E HISTOLOGIA DENTAL PDFDocumento198 páginasLeccion 2. EMBRIOLOGIA E HISTOLOGIA DENTAL PDFEmmanuel CieloAún no hay calificaciones
- Trabajo Fin de Master AzidiolDocumento89 páginasTrabajo Fin de Master Azidiolaguadepila100% (1)
- TOXICOLOGIADocumento12 páginasTOXICOLOGIAChelythaAún no hay calificaciones
- LaringitisDocumento1 páginaLaringitiscapa correoAún no hay calificaciones
- Monografía - TuberculosisDocumento22 páginasMonografía - TuberculosisBilly Solano Jesus100% (2)
- Resumen Nariz y BocaDocumento38 páginasResumen Nariz y BocaJose SantanaAún no hay calificaciones
- Medicina III - Trastornos Digestivos FuncionalesDocumento29 páginasMedicina III - Trastornos Digestivos FuncionalesUSMP FN ARCHIVOS100% (1)
- Quemaduras ElectricasDocumento42 páginasQuemaduras ElectricasJohn ChaucaAún no hay calificaciones
- Historia Natural de La EnfermedadDocumento21 páginasHistoria Natural de La EnfermedadCristopher Barboza GarciaAún no hay calificaciones
- Amala y KamalaDocumento1 páginaAmala y KamalaJOHANA CARMONA VALDIVIAAún no hay calificaciones
- FLP VenezuelaDocumento2 páginasFLP VenezuelaRafael ZambranoAún no hay calificaciones
- Atelectasia PulmonarDocumento79 páginasAtelectasia PulmonarFabri CarbajalAún no hay calificaciones
- Columna vertebral: estructura, funciones y alteracionesDocumento40 páginasColumna vertebral: estructura, funciones y alteracionesDarkzz KagamineAún no hay calificaciones
- Medula Espinal Anatomofisiologia AsgDocumento18 páginasMedula Espinal Anatomofisiologia AsgHumberto CarvajalAún no hay calificaciones
- Curar linfadenitis en cuyes: causas, síntomas y tratamientoDocumento4 páginasCurar linfadenitis en cuyes: causas, síntomas y tratamientoDaniel Taca100% (1)
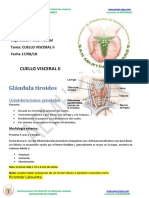
- Degrabada de Glandula Tiroides-Aemh2018 PDFDocumento6 páginasDegrabada de Glandula Tiroides-Aemh2018 PDFLuis TapiaAún no hay calificaciones
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Notas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesDe EverandNotas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesAún no hay calificaciones
- TDAH en Adultos. Cómo Reconocer y Tratar a un Adulto con TDAH en 30 Fáciles PasosDe EverandTDAH en Adultos. Cómo Reconocer y Tratar a un Adulto con TDAH en 30 Fáciles PasosCalificación: 4 de 5 estrellas4/5 (8)
- Prescripción de ejercico físico para la saludDe EverandPrescripción de ejercico físico para la saludCalificación: 5 de 5 estrellas5/5 (1)
- Fisiopatología de las enfermedades cardiovascularesDe EverandFisiopatología de las enfermedades cardiovascularesCalificación: 5 de 5 estrellas5/5 (1)
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- La metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceDe EverandLa metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceCalificación: 5 de 5 estrellas5/5 (8)
- Terapia cognitiva: Conceptos básicos y profundizaciónDe EverandTerapia cognitiva: Conceptos básicos y profundizaciónCalificación: 5 de 5 estrellas5/5 (1)
- El libro conciso de la punción seca: Manual del terapeuta para las aplicaciones en los puntos gatillo miofasciales (Color)De EverandEl libro conciso de la punción seca: Manual del terapeuta para las aplicaciones en los puntos gatillo miofasciales (Color)Calificación: 3 de 5 estrellas3/5 (2)
- Genética general: Libro de textoDe EverandGenética general: Libro de textoCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Neuropsicología: Los fundamentos de la materiaDe EverandNeuropsicología: Los fundamentos de la materiaCalificación: 5 de 5 estrellas5/5 (1)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- Dieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasDe EverandDieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasCalificación: 4 de 5 estrellas4/5 (9)
- La Dieta Mediterránea Para Principiantes, Guía Paso A Paso Con Recetas Para Comer Mejor Y Bajar De PesoDe EverandLa Dieta Mediterránea Para Principiantes, Guía Paso A Paso Con Recetas Para Comer Mejor Y Bajar De PesoCalificación: 5 de 5 estrellas5/5 (2)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- Cerebros rotos: Pacientes asombrosos que me enseñaron a vivirDe EverandCerebros rotos: Pacientes asombrosos que me enseñaron a vivirCalificación: 5 de 5 estrellas5/5 (3)
- Nuestra mente nos engaña: Sesgos y errores cognitivos que todos cometemosDe EverandNuestra mente nos engaña: Sesgos y errores cognitivos que todos cometemosCalificación: 3.5 de 5 estrellas3.5/5 (6)
- Resumen de Pensar rápido pensar despacio de Daniel KahnemanDe EverandResumen de Pensar rápido pensar despacio de Daniel KahnemanCalificación: 4.5 de 5 estrellas4.5/5 (11)
- El cerebro del niño explicado a los padresDe EverandEl cerebro del niño explicado a los padresCalificación: 4.5 de 5 estrellas4.5/5 (147)
- Sana tu Cuerpo, Calma tu Mente: Desintoxicar Hígado, Intestino Permeable, Salud Hormonal, Curación Emocional, Relajación, Ansiedad y Sanidad mental, Atención Plena, Psicoterapia y NutriciónDe EverandSana tu Cuerpo, Calma tu Mente: Desintoxicar Hígado, Intestino Permeable, Salud Hormonal, Curación Emocional, Relajación, Ansiedad y Sanidad mental, Atención Plena, Psicoterapia y NutriciónCalificación: 5 de 5 estrellas5/5 (4)
- Psiconeuroinmunología para la práctica clínicaDe EverandPsiconeuroinmunología para la práctica clínicaCalificación: 5 de 5 estrellas5/5 (4)
- 200 tareas en terapia breve: 2ª ediciónDe Everand200 tareas en terapia breve: 2ª ediciónCalificación: 4.5 de 5 estrellas4.5/5 (33)
- Psicoterapia breve con niños y adolescentesDe EverandPsicoterapia breve con niños y adolescentesCalificación: 4.5 de 5 estrellas4.5/5 (15)
- Abrázame fuerte: Siete conversaciones para un amor duraderoDe EverandAbrázame fuerte: Siete conversaciones para un amor duraderoManu BerásteguiCalificación: 4.5 de 5 estrellas4.5/5 (13)
- Psicópatas seriales: Un recorrido por su oscura e inquietante naturalezaDe EverandPsicópatas seriales: Un recorrido por su oscura e inquietante naturalezaCalificación: 4 de 5 estrellas4/5 (3)
- Los cinco ritos tibetanos de la eterna juventud: El ojo de la revelaciónDe EverandLos cinco ritos tibetanos de la eterna juventud: El ojo de la revelaciónCalificación: 4.5 de 5 estrellas4.5/5 (12)