También podría gustarte
- Generalidades Del MetabolismoDocumento12 páginasGeneralidades Del MetabolismoMaria Paula GarciaAún no hay calificaciones
- La Ciencia Del Metabolismo: Metabolismo sin rodeosDe EverandLa Ciencia Del Metabolismo: Metabolismo sin rodeosCalificación: 5 de 5 estrellas5/5 (1)
- Cuestionario BQMDocumento4 páginasCuestionario BQMBeatriz G.Aún no hay calificaciones
- Ayudas ergogénicas y nutricionalesDe EverandAyudas ergogénicas y nutricionalesCalificación: 5 de 5 estrellas5/5 (2)
- CARBOHIDRATOS UNIFICADOoDocumento11 páginasCARBOHIDRATOS UNIFICADOoJesus Smaikel Teran MuenalaAún no hay calificaciones
- Nutrición: La solución para vivir sin enfermedadesDe EverandNutrición: La solución para vivir sin enfermedadesAún no hay calificaciones
- Tema 1 - INTRODUCCIÓN AL METABOLISMODocumento7 páginasTema 1 - INTRODUCCIÓN AL METABOLISMOvalerialarrarte01Aún no hay calificaciones
- Integración y Regulación Metabólica - Resumen Qca IIDocumento15 páginasIntegración y Regulación Metabólica - Resumen Qca IIVEROAún no hay calificaciones
- Vias Que Procesan Los Principales Productos de La DigestionDocumento6 páginasVias Que Procesan Los Principales Productos de La DigestionJulietta CondeñaAún no hay calificaciones
- Clase 12-Nutricion, Metabolismo, Ciclo Del Acido Citrico, Glucolisis 2018Documento52 páginasClase 12-Nutricion, Metabolismo, Ciclo Del Acido Citrico, Glucolisis 2018Natalia0% (1)
- Nbnu - U1 - Ea - KavdDocumento11 páginasNbnu - U1 - Ea - KavdKarina De La PazAún no hay calificaciones
- Taller Segundo CorteDocumento8 páginasTaller Segundo CorteGustavo AdolfoAún no hay calificaciones
- Las Vías Metabólicas Pueden Estudiarse A Diferentes Niveles de OrganizaciónDocumento4 páginasLas Vías Metabólicas Pueden Estudiarse A Diferentes Niveles de OrganizaciónRikelbin PujolsAún no hay calificaciones
- Respuesta Metabolica Durante El Ayuno La Sepsis y La InjuriDocumento30 páginasRespuesta Metabolica Durante El Ayuno La Sepsis y La InjuriMTNC. César ZermeñoAún no hay calificaciones
- Integracion MetabolicaDocumento21 páginasIntegracion Metabolicajuan100% (2)
- Metabolismo CHO, Lípidos y ProteínasDocumento23 páginasMetabolismo CHO, Lípidos y ProteínasAdrian MorganAún no hay calificaciones
- Metabolismo de CarbohidratosDocumento4 páginasMetabolismo de Carbohidratospao187Aún no hay calificaciones
- LA ALIMENTACION Y NUTRICION EN POLIGASTRICOS Examen 2Documento17 páginasLA ALIMENTACION Y NUTRICION EN POLIGASTRICOS Examen 2ManuelAún no hay calificaciones
- Metabolismo de Principales Alimentos 2023Documento17 páginasMetabolismo de Principales Alimentos 2023Arnoldo Jose Candanedo JaramilloAún no hay calificaciones
- Importancia Del Metabolismo BasalDocumento7 páginasImportancia Del Metabolismo BasalIsrael Grover Condorcet MancillaAún no hay calificaciones
- PARTE 2 Metabolismos - ABIDocumento66 páginasPARTE 2 Metabolismos - ABInm5611645Aún no hay calificaciones
- Práctica1 Cuestionario FernandezDocumento6 páginasPráctica1 Cuestionario Fernandezyadira fernandezAún no hay calificaciones
- TEMA 1 MetabolismoDocumento14 páginasTEMA 1 MetabolismoBerenize VidalAún no hay calificaciones
- Guia de Integracion Metabolica CompletaDocumento19 páginasGuia de Integracion Metabolica CompletaBrianAún no hay calificaciones
- 02 PTCourseWeek1 - Bioquimica 101Documento13 páginas02 PTCourseWeek1 - Bioquimica 101El SandroAún no hay calificaciones
- Rutas MetabolicasDocumento3 páginasRutas Metabolicasjose luis de la cruzAún no hay calificaciones
- Seminario. Glucolisis y Oxidacion Del Piruvato.Documento45 páginasSeminario. Glucolisis y Oxidacion Del Piruvato.lorena navarreteAún no hay calificaciones
- Metabolismo Leidy Tatiana AcevedoDocumento16 páginasMetabolismo Leidy Tatiana AcevedoÄcëvëdö TatianaAún no hay calificaciones
- Vías Que Procesan Los Principales Productos de La DigestiónDocumento6 páginasVías Que Procesan Los Principales Productos de La DigestiónROSA PORTERO RODRIGUEZAún no hay calificaciones
- Ensayo PropeDocumento14 páginasEnsayo PropeFenanda mar redondoAún no hay calificaciones
- TEMA 02 - Metabolismo de Tejidos y ÓrganosDocumento16 páginasTEMA 02 - Metabolismo de Tejidos y ÓrganosDavid García MartínezAún no hay calificaciones
- Metabolismo Actividades - DiazDocumento14 páginasMetabolismo Actividades - Diazaildy27bascaraAún no hay calificaciones
- Resumen FinalDocumento8 páginasResumen Finalsusan uribeAún no hay calificaciones
- DriveDocumento16 páginasDriveLau Rafael Flores RiveraAún no hay calificaciones
- Bioqimica AntologiaDocumento9 páginasBioqimica AntologiaIsrael Dzul LopezAún no hay calificaciones
- CB Carbohidratos ResidPeru 11Documento9 páginasCB Carbohidratos ResidPeru 11Alex Cárdenas ValdiviesoAún no hay calificaciones
- Bioquimica Resumen H 601512 Downloadable 3597272Documento14 páginasBioquimica Resumen H 601512 Downloadable 3597272Fabián OlivaresAún no hay calificaciones
- Balotario Cap LipidosDocumento6 páginasBalotario Cap LipidosdiaconalAún no hay calificaciones
- Harper - Generalidades Del MetabolismoDocumento12 páginasHarper - Generalidades Del MetabolismoSamuel Morales NavarroAún no hay calificaciones
- Tema 16Documento7 páginasTema 16IvanaAún no hay calificaciones
- Integracion Metabolica1Documento26 páginasIntegracion Metabolica1Isaac EspinozaAún no hay calificaciones
- Metabolismo de Los GlúcidosDocumento10 páginasMetabolismo de Los GlúcidosAguus CabreraAún no hay calificaciones
- Metabolismo y Equilibrio EnergeticoDocumento31 páginasMetabolismo y Equilibrio Energeticoshirley andradeAún no hay calificaciones
- Introduccion Ensayo 10Documento7 páginasIntroduccion Ensayo 10santiagoAún no hay calificaciones
- Ciclo de KrebsDocumento3 páginasCiclo de KrebsNatalia JiménezAún no hay calificaciones
- Bio Qui MicaDocumento21 páginasBio Qui Micalefef81847Aún no hay calificaciones
- Iefi NutriciónDocumento14 páginasIefi NutriciónAgustina DaricoAún no hay calificaciones
- Bioquimica Material DigDocumento179 páginasBioquimica Material DigMonica RodriguezAún no hay calificaciones
- Taller # 1 CarbohidratosDocumento10 páginasTaller # 1 CarbohidratossaraAún no hay calificaciones
- Practica 8 METABOLISMODocumento12 páginasPractica 8 METABOLISMOSimon Jesus Gonzalez ArangoAún no hay calificaciones
- Metabolismo EnergeticoDocumento21 páginasMetabolismo EnergeticonataliamaritzaladinoAún no hay calificaciones
- CarboDocumento12 páginasCarboLina QuinteroAún no hay calificaciones
- TALLER - No. - 1 - de - BIOQUIMICA - 2021 - 2 - DEFDocumento28 páginasTALLER - No. - 1 - de - BIOQUIMICA - 2021 - 2 - DEFlaura pardoAún no hay calificaciones
- Metabolismo - Química Biológica IDocumento7 páginasMetabolismo - Química Biológica ILuján CarossiniAún no hay calificaciones
- Unidad V. Tema 5. Carbohidratos, GlicolisisDocumento16 páginasUnidad V. Tema 5. Carbohidratos, GlicolisisGlenda G. OrtizAún no hay calificaciones
- Interacciones MetabólicasDocumento33 páginasInteracciones MetabólicasAugusto Suárez Gómez100% (3)
- Aporte - Trabajo Colaborativo - Bioquímica - 2016-2Documento8 páginasAporte - Trabajo Colaborativo - Bioquímica - 2016-2Jeffe Yepes GarcíaAún no hay calificaciones
- Biosisntesis de CarbohidratosDocumento5 páginasBiosisntesis de Carbohidratostiburcio floresAún no hay calificaciones
- Actividad 9 - Integración Metabólica 1Documento21 páginasActividad 9 - Integración Metabólica 1Tomas VillanuevaAún no hay calificaciones
- Esclerosis SistemicaDocumento26 páginasEsclerosis SistemicaChemicalero FabulosoaguafiestaAún no hay calificaciones
- Estudio Mediante Tecnicas Inmunoquimicas Del Efecto Del Procesado en Algunas Proteina AlergenicasDocumento210 páginasEstudio Mediante Tecnicas Inmunoquimicas Del Efecto Del Procesado en Algunas Proteina AlergenicasKiko SalvadorAún no hay calificaciones
- Unidad 2 - Sistema DigestivoDocumento15 páginasUnidad 2 - Sistema DigestivoNeri MercedesAún no hay calificaciones
- Resumen Páncreas Finn GeneserDocumento3 páginasResumen Páncreas Finn GeneserLaura FMAún no hay calificaciones
- GasesDocumento6 páginasGasesWildor CastilloAún no hay calificaciones
- Lanchas en La Bahia FinalDocumento23 páginasLanchas en La Bahia FinaljavierAún no hay calificaciones
- TP 0 Ciclo Sexual Resuelto1 FSH UNLAMDocumento8 páginasTP 0 Ciclo Sexual Resuelto1 FSH UNLAMDayannelopes123Aún no hay calificaciones
- Enfermedad de Hirschsprung o Megacolon Agangliónico CongénitoDocumento14 páginasEnfermedad de Hirschsprung o Megacolon Agangliónico CongénitoyamileAún no hay calificaciones
- copia.docxDocumento67 páginascopia.docxSebastian Hernandez UknowAún no hay calificaciones
- TuberculosisDocumento19 páginasTuberculosisapi-3814371100% (2)
- Castigos de La AntiguedadDocumento7 páginasCastigos de La AntiguedadMiller Alvarado CarretoAún no hay calificaciones
- Fundamentos de Ornitologia y AviturismoDocumento25 páginasFundamentos de Ornitologia y AviturismoPanchin David0% (1)
- Presion OsmoticaDocumento6 páginasPresion OsmoticaWilson Piero Fernández VelaAún no hay calificaciones
- Accesos Vasculares PDFDocumento53 páginasAccesos Vasculares PDFEukary DiazAún no hay calificaciones
- Neuropsicologia de La Conducta SexualDocumento22 páginasNeuropsicologia de La Conducta SexualLuis Stanley SalinasAún no hay calificaciones
- Embriologia Del Aparato RespiratorioDocumento11 páginasEmbriologia Del Aparato RespiratorioMartín Inmediato Ghetti100% (1)
- Rehabilitacion BrainportDocumento24 páginasRehabilitacion BrainportgalenoredAún no hay calificaciones
- Aparo UrinarioDocumento7 páginasAparo UrinarioRonal Lizana SantosAún no hay calificaciones
- Vías de Administracion de MedicamentosDocumento10 páginasVías de Administracion de Medicamentosharoldveramarin100% (11)
- Bioelemento y BiomoleculaDocumento8 páginasBioelemento y BiomoleculaRoyman BernalAún no hay calificaciones
- Diabetes Insípida NefrogénicaDocumento15 páginasDiabetes Insípida NefrogénicaEsme Tecuanhuehue PaulinoAún no hay calificaciones
- Tipos AnemiaDocumento154 páginasTipos AnemiaTetsuo ShimaAún no hay calificaciones
- 2do Grado-Semana 19Documento3 páginas2do Grado-Semana 19Geydy TaipeAún no hay calificaciones
- Farmacología MolecularDocumento8 páginasFarmacología MolecularEduardo Dávila100% (2)
- Guía de Actividades BioquímicaDocumento5 páginasGuía de Actividades BioquímicaMary PerezAún no hay calificaciones
- Cómo Detectar MentirasDocumento6 páginasCómo Detectar MentirasJosé ManuelAún no hay calificaciones
- 3er Año Biología Heft Weiss 2Documento15 páginas3er Año Biología Heft Weiss 2xiomara cayroAún no hay calificaciones
- Informe 5 BCTII Endocrinologia Del EembarazoDocumento14 páginasInforme 5 BCTII Endocrinologia Del EembarazoIanCurtisAún no hay calificaciones
- Qué Es Una Angina de PechoDocumento5 páginasQué Es Una Angina de PechoDaniel Lévano AlzamoraAún no hay calificaciones
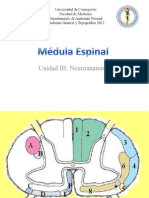
- Médula Espinal - Vías AscendentesDocumento47 páginasMédula Espinal - Vías AscendentesNicolás Alejandro Peña Toro90% (21)
- Homo antecessor: El nacimiento de una especieDe EverandHomo antecessor: El nacimiento de una especieCalificación: 5 de 5 estrellas5/5 (1)
- DIETA MEDITERRANEA - Mejores Recetas de la Cocina Mediterranea Para Bajar de Peso SaludablementeDe EverandDIETA MEDITERRANEA - Mejores Recetas de la Cocina Mediterranea Para Bajar de Peso SaludablementeCalificación: 5 de 5 estrellas5/5 (4)
- GuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisDe EverandGuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisCalificación: 4 de 5 estrellas4/5 (9)
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- Notas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesDe EverandNotas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesAún no hay calificaciones
- Sana tu Cuerpo, Calma tu Mente: Desintoxicar Hígado, Intestino Permeable, Salud Hormonal, Curación Emocional, Relajación, Ansiedad y Sanidad mental, Atención Plena, Psicoterapia y NutriciónDe EverandSana tu Cuerpo, Calma tu Mente: Desintoxicar Hígado, Intestino Permeable, Salud Hormonal, Curación Emocional, Relajación, Ansiedad y Sanidad mental, Atención Plena, Psicoterapia y NutriciónCalificación: 5 de 5 estrellas5/5 (4)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Alimentación antiinflamatoria: Primeros pasos para poner en práctica una alimentación saludableDe EverandAlimentación antiinflamatoria: Primeros pasos para poner en práctica una alimentación saludableCalificación: 4.5 de 5 estrellas4.5/5 (119)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- Neuropsicología: Los fundamentos de la materiaDe EverandNeuropsicología: Los fundamentos de la materiaCalificación: 5 de 5 estrellas5/5 (1)
- Mente sana, cuerpo sano: El poder de una mentalidad saludableDe EverandMente sana, cuerpo sano: El poder de una mentalidad saludableCalificación: 5 de 5 estrellas5/5 (90)
- Postres Saludables para Disfrutar sin Culpa. Postres Paleo sin Gluten, Azucar ni LactosaDe EverandPostres Saludables para Disfrutar sin Culpa. Postres Paleo sin Gluten, Azucar ni LactosaCalificación: 5 de 5 estrellas5/5 (17)
- La Dieta Mediterránea Para Principiantes, Guía Paso A Paso Con Recetas Para Comer Mejor Y Bajar De PesoDe EverandLa Dieta Mediterránea Para Principiantes, Guía Paso A Paso Con Recetas Para Comer Mejor Y Bajar De PesoCalificación: 5 de 5 estrellas5/5 (2)
- La Dieta Antiinflamatoria: Haz estos cambios simples y económicos en tu dieta y comienza a sentirte mejor dentro de 24 horas! (Spanish Edition)De EverandLa Dieta Antiinflamatoria: Haz estos cambios simples y económicos en tu dieta y comienza a sentirte mejor dentro de 24 horas! (Spanish Edition)Calificación: 5 de 5 estrellas5/5 (4)
- Nuestra mente nos engaña: Sesgos y errores cognitivos que todos cometemosDe EverandNuestra mente nos engaña: Sesgos y errores cognitivos que todos cometemosCalificación: 3.5 de 5 estrellas3.5/5 (6)
- Minerales y vitaminas: Micronutrientes esenciales en la alimentación, nutrición y salud.De EverandMinerales y vitaminas: Micronutrientes esenciales en la alimentación, nutrición y salud.Calificación: 3 de 5 estrellas3/5 (2)
- Altas Dosis: Los efectos milagrosos de dosis extremadamente altas de vitamina D3. El gran secreto que la industria farmacéutica quiere ocultarle.De EverandAltas Dosis: Los efectos milagrosos de dosis extremadamente altas de vitamina D3. El gran secreto que la industria farmacéutica quiere ocultarle.Calificación: 5 de 5 estrellas5/5 (4)
- Sacale partido a tu cerebro: Todo lo que necesitas saber para mejorar tu memoria, tomar decisiones y aprovechar todo tu potencialDe EverandSacale partido a tu cerebro: Todo lo que necesitas saber para mejorar tu memoria, tomar decisiones y aprovechar todo tu potencialCalificación: 3 de 5 estrellas3/5 (1)
- El cerebro del niño explicado a los padresDe EverandEl cerebro del niño explicado a los padresCalificación: 4.5 de 5 estrellas4.5/5 (147)
- Cerebros rotos: Pacientes asombrosos que me enseñaron a vivirDe EverandCerebros rotos: Pacientes asombrosos que me enseñaron a vivirCalificación: 5 de 5 estrellas5/5 (3)
- Libro de cocina completo para diabéticos en español/ Diabetic cookbook in spanishDe EverandLibro de cocina completo para diabéticos en español/ Diabetic cookbook in spanishCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Autofagia: Descubra los Secretos para la Pérdida de Peso, el Rejuvenecimiento y la Curación con el Ayuno Intermitente y ProlongadoDe EverandAutofagia: Descubra los Secretos para la Pérdida de Peso, el Rejuvenecimiento y la Curación con el Ayuno Intermitente y ProlongadoCalificación: 4 de 5 estrellas4/5 (15)