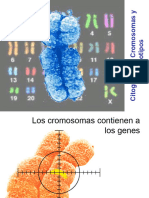
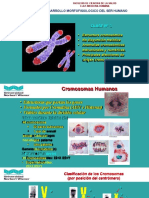
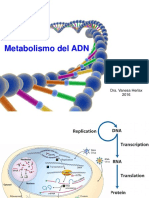
También podría gustarte
- Clase 7 Ciclo Celular MitosisDocumento34 páginasClase 7 Ciclo Celular MitosisAlejandro Rivera Meneses100% (1)
- Para Finalizar - Biología - 2do B.G.U - Página 74,75Documento6 páginasPara Finalizar - Biología - 2do B.G.U - Página 74,75Anonymous CAMGgn85% (186)
- Introducción a la Biología: RESÚMENES UNIVERSITARIOSDe EverandIntroducción a la Biología: RESÚMENES UNIVERSITARIOSCalificación: 5 de 5 estrellas5/5 (1)
- Efecto Letal Y Mutagenico de La Radiacion Ultravioleta Sobre Los MicroorganismosDocumento27 páginasEfecto Letal Y Mutagenico de La Radiacion Ultravioleta Sobre Los MicroorganismosAntony LanderosAún no hay calificaciones
- Clase 3.cromatinaDocumento82 páginasClase 3.cromatinaMaria Victoria Torres Lara100% (1)
- Mecanismos de Reparación Del ADNDocumento7 páginasMecanismos de Reparación Del ADNVIOLETA NICOLE LINARES PORRASAún no hay calificaciones
- Biologia Celular - Clase 11Documento47 páginasBiologia Celular - Clase 11api-3701781Aún no hay calificaciones
- Compartimientos Celulares: Núcleo y Citoplasma: Biología Celular y Del DesarrolloDocumento98 páginasCompartimientos Celulares: Núcleo y Citoplasma: Biología Celular y Del DesarrolloGonzalo VaroneAún no hay calificaciones
- 12 - Power BiologiaDocumento31 páginas12 - Power BiologiaLuis Armando PandoAún no hay calificaciones
- Núcleo CelularDocumento17 páginasNúcleo CelularNorman NeyraAún no hay calificaciones
- Clase 3. Mecanismos Geneticos Básicos I 2021Documento65 páginasClase 3. Mecanismos Geneticos Básicos I 2021María Belén MuñozAún no hay calificaciones
- Semana 7 Núcleo BIOCEL MEHU 2021 10Documento38 páginasSemana 7 Núcleo BIOCEL MEHU 2021 10Ivan Vargas LopezAún no hay calificaciones
- Asm Bi1 TS009Documento15 páginasAsm Bi1 TS009DLEO IAún no hay calificaciones
- BIOL-034 Clase 3-Mecanismos Genéticos Básicos IDocumento32 páginasBIOL-034 Clase 3-Mecanismos Genéticos Básicos Ibenjamin pinilla rojasAún no hay calificaciones
- Tema3 - Compartimento NuclearDocumento40 páginasTema3 - Compartimento NuclearCristina Hidalgo RivasAún no hay calificaciones
- Núcleo Celular: Colegio Mercedario "San Pedro Pascual"Documento5 páginasNúcleo Celular: Colegio Mercedario "San Pedro Pascual"William ChuraAún no hay calificaciones
- Diagramas - GenéticaDocumento12 páginasDiagramas - GenéticaFrida MagaliAún no hay calificaciones
- 3-04 CitogenéticaDocumento56 páginas3-04 Citogenéticamaria jose millan albornozAún no hay calificaciones
- Plantilla Clase T 3 DMFSHDocumento40 páginasPlantilla Clase T 3 DMFSHMarylin SanchezAún no hay calificaciones
- Plenaria Envoltura Nuclear y Replicación - Septiembre 26Documento24 páginasPlenaria Envoltura Nuclear y Replicación - Septiembre 26Estefanía HerreraAún no hay calificaciones
- Unidad 4 - Parte 2 de 6Documento15 páginasUnidad 4 - Parte 2 de 6Jose FranciscoAún no hay calificaciones
- 2° Parcial Biología - PDF?Documento35 páginas2° Parcial Biología - PDF?tatyy7177Aún no hay calificaciones
- Unidad V - Nucleo y División CelularDocumento109 páginasUnidad V - Nucleo y División CelularJennifer RodríguezAún no hay calificaciones
- Nucleo y Nucleolo PDFDocumento11 páginasNucleo y Nucleolo PDFRellir830YTAún no hay calificaciones
- Solemne 3 BiologíaDocumento24 páginasSolemne 3 BiologíakarlarenatavalenzuelasAún no hay calificaciones
- NúcleoDocumento1 páginaNúcleoMariana Guerrero TurrubiatesAún no hay calificaciones
- Núcleo: InterfaseDocumento51 páginasNúcleo: InterfaseHernán Hernández MarcialAún no hay calificaciones
- Genoma Humano 1Documento47 páginasGenoma Humano 1Noelia RiqueroAún no hay calificaciones
- 1 - Material GeneticoDocumento45 páginas1 - Material GeneticoCatalina Sotomayor GutiérrezAún no hay calificaciones
- Nucleo CelularDocumento64 páginasNucleo CelularCib3r V1pAún no hay calificaciones
- T2 - Cromatina, Cromosomas y 1ra Ley Mendel 2015Documento44 páginasT2 - Cromatina, Cromosomas y 1ra Ley Mendel 2015Gabriela GarcíaAún no hay calificaciones
- I.bloque Transmision Y Expresion de La Informacion Genetica: 1. Tema. IntroduccionDocumento48 páginasI.bloque Transmision Y Expresion de La Informacion Genetica: 1. Tema. IntroduccionepepAún no hay calificaciones
- S9 - NucleoDocumento9 páginasS9 - NucleoJhony Farith Farfán ReáteguiAún no hay calificaciones
- Clase 14. DNA y ReplicaciónDocumento39 páginasClase 14. DNA y ReplicaciónaylinesotorAún no hay calificaciones
- Clase Teorica 2 PDFDocumento42 páginasClase Teorica 2 PDFLuis PeredoAún no hay calificaciones
- Replicacion Del Adn 2013 14Documento48 páginasReplicacion Del Adn 2013 14YaiAún no hay calificaciones
- Tema 4. Citología. Nucleo y Nucleolo. RibosomasDocumento24 páginasTema 4. Citología. Nucleo y Nucleolo. RibosomasjanynedosAún no hay calificaciones
- Biología - Ciclo Celular - Semestral SMDocumento13 páginasBiología - Ciclo Celular - Semestral SMBeltran Remigio Champi NoaAún no hay calificaciones
- C2. ADN y Ciclo CelularDocumento22 páginasC2. ADN y Ciclo CelularMaria Jesus MascayanoAún no hay calificaciones
- Condensación Del DNA y CromosomasDocumento46 páginasCondensación Del DNA y CromosomasManuela Hoyos GAún no hay calificaciones
- Dna, Genes y CromosomasDocumento4 páginasDna, Genes y CromosomasXimena Duran AyalaAún no hay calificaciones
- Clase 3 Organizacion Del Dna en Cromosomas - 1863 - 0Documento44 páginasClase 3 Organizacion Del Dna en Cromosomas - 1863 - 0Rebeca RojasAún no hay calificaciones
- 3ra Clase - Estructura CromosomicaDocumento41 páginas3ra Clase - Estructura CromosomicaOliver Manrique MaldonadoAún no hay calificaciones
- ADN y Reproducción CelularDocumento81 páginasADN y Reproducción CelularJorgeAún no hay calificaciones
- Z-Tema 2 Biol Mol ADNDocumento12 páginasZ-Tema 2 Biol Mol ADNLuz Zapata PardoAún no hay calificaciones
- Clase NucleoDocumento43 páginasClase NucleoElizabeth Trinidad RuizAún no hay calificaciones
- Cloroplasto y NúcleoDocumento6 páginasCloroplasto y NúcleoFernanda UribeAún no hay calificaciones
- Apuntes Biología 3Documento37 páginasApuntes Biología 3mbelenruhlAún no hay calificaciones
- Nucleo Celular - Replicacion 2021-IIDocumento28 páginasNucleo Celular - Replicacion 2021-IIKaren FelixAún no hay calificaciones
- Tema 28 CromosomasDocumento15 páginasTema 28 CromosomasVanessa PeñaAún no hay calificaciones
- 03 Organizacion Cromatina2021Documento35 páginas03 Organizacion Cromatina2021tavogreAún no hay calificaciones
- Presentación Biología Organización Del Ácido Desoxirribonucleico (ADN) Ilustrativo AzulDocumento13 páginasPresentación Biología Organización Del Ácido Desoxirribonucleico (ADN) Ilustrativo AzulsomontalbancaAún no hay calificaciones
- Wuolah Free Genetica VeterinariaDocumento37 páginasWuolah Free Genetica VeterinariafisiovetvigoAún no hay calificaciones
- Solemne 1 ADN, Cromosomas, Replicacion y Reparacion Del ADNDocumento24 páginasSolemne 1 ADN, Cromosomas, Replicacion y Reparacion Del ADNBasthian CarvalloAún no hay calificaciones
- 3ra Clase - DR ARCOS Cromosomas Humanos Anomalías Cromosómicas Numéricas y EstructuralesDocumento50 páginas3ra Clase - DR ARCOS Cromosomas Humanos Anomalías Cromosómicas Numéricas y Estructuralesalexis yupanquiAún no hay calificaciones
- Tema 10Documento43 páginasTema 10xXShadow.J XxAún no hay calificaciones
- Mapa NucleoDocumento1 páginaMapa NucleoLuis Eduardo de Paz BarreraAún no hay calificaciones
- Clase 2 TM2017 Núcleo y CromosomasDocumento52 páginasClase 2 TM2017 Núcleo y CromosomasVale NtinaAún no hay calificaciones
- Tema 8 El Ciclo Celular Eucariota IDocumento30 páginasTema 8 El Ciclo Celular Eucariota ISplit GamesAún no hay calificaciones
- Biología EfiDocumento30 páginasBiología EfiJuuAún no hay calificaciones
- DMFSH CT 3Documento48 páginasDMFSH CT 3anamarAún no hay calificaciones
- Resumen de Bio Uba Xxi 2ºpDocumento26 páginasResumen de Bio Uba Xxi 2ºpFiorella GurziAún no hay calificaciones
- Tema 3 - Herramientas para La Modificacion Dirigida Del Genoma APUNTESDocumento12 páginasTema 3 - Herramientas para La Modificacion Dirigida Del Genoma APUNTESAndrea EsquivelAún no hay calificaciones
- Mutación, Recombinación Genética y Flugo GénicoDocumento36 páginasMutación, Recombinación Genética y Flugo GénicoJair elias SanchezAún no hay calificaciones
- Replicacion de AdnDocumento2 páginasReplicacion de AdnSofiaAún no hay calificaciones
- Encuadre Bioquímica Médica 2022-2 301 y 302Documento4 páginasEncuadre Bioquímica Médica 2022-2 301 y 302David Morachis OñateAún no hay calificaciones
- MFPH IiDocumento57 páginasMFPH IiJesshia LugoAún no hay calificaciones
- REPARACIÓNDocumento3 páginasREPARACIÓNJoseph Mena VidalAún no hay calificaciones
- Reproducción: Generación de Ganado Modificado Genéticamente Mediante Transferencia Nuclear de Células SomáticasDocumento12 páginasReproducción: Generación de Ganado Modificado Genéticamente Mediante Transferencia Nuclear de Células SomáticasAdolfo Mora SanchezAún no hay calificaciones
- Biologia BancoDocumento3 páginasBiologia BancoJeniffer MedinaAún no hay calificaciones
- Ataxia TelangectasiaDocumento8 páginasAtaxia TelangectasiaMaximilianoAún no hay calificaciones
- Reparación Del AdnDocumento14 páginasReparación Del AdnAndres Salazar100% (1)
- MIT Biologia MolecularDocumento8 páginasMIT Biologia MolecularJR Bio GenAún no hay calificaciones
- Seminario Metabolismo ADN 2016Documento51 páginasSeminario Metabolismo ADN 2016Melanie TassoAún no hay calificaciones
- Tipos Replicacion Reparacion de AdnDocumento83 páginasTipos Replicacion Reparacion de AdnCristopher Pinedo CarranzaAún no hay calificaciones
- El ADN Bajo El Efecto Del SolDocumento5 páginasEl ADN Bajo El Efecto Del SolDafne Hirlani Rodriguez GonzalezAún no hay calificaciones
- CAPÍTULO 476 - Biología Del EnvejecimientoDocumento13 páginasCAPÍTULO 476 - Biología Del EnvejecimientoAlvaro IbarraAún no hay calificaciones
- Unidad 2 Mantenimiento Correctivo Modulo 1Documento45 páginasUnidad 2 Mantenimiento Correctivo Modulo 1Nicholas SmithAún no hay calificaciones
- Bmi TDocumento176 páginasBmi TJuan Arcos HuasasquicheAún no hay calificaciones
- Guía Biología MolecularDocumento25 páginasGuía Biología MolecularJay GarrickAún no hay calificaciones
- Copia de GENETICADocumento15 páginasCopia de GENETICAPaulina SánchezAún no hay calificaciones
- Cancerogenesis Tarea1111111111Documento14 páginasCancerogenesis Tarea1111111111Cecilia Nurmy Yareli Cecilio EnriquezAún no hay calificaciones
- Trabajo Practico MutacionesDocumento3 páginasTrabajo Practico Mutacionesbian rotelaAún no hay calificaciones
- GeneticaDocumento11 páginasGeneticaKarla MendozaAún no hay calificaciones
- Sesión 6 - Recombinación GenéticaDocumento14 páginasSesión 6 - Recombinación GenéticaVale HAún no hay calificaciones
- Vias de Reparacion Del DNADocumento7 páginasVias de Reparacion Del DNAbrailercordobaAún no hay calificaciones
- Biofísica BCM ResumenDocumento17 páginasBiofísica BCM ResumenAriani PrestesAún no hay calificaciones
- Primer Tema Patologia-FusionadoDocumento54 páginasPrimer Tema Patologia-FusionadoSofia ParedezAún no hay calificaciones
- Unidad 2 Mantenimiento CorrectivoDocumento40 páginasUnidad 2 Mantenimiento CorrectivoNicholas SmithAún no hay calificaciones