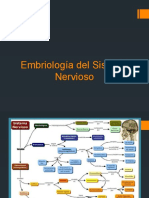
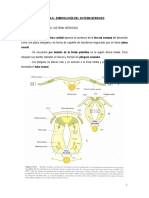
También podría gustarte
- Histología y Embrología del Sitema NerviosoDe EverandHistología y Embrología del Sitema NerviosoCalificación: 4 de 5 estrellas4/5 (4)
- Anatomía y Fisiología del Sistema NerviosoDe EverandAnatomía y Fisiología del Sistema NerviosoCalificación: 4 de 5 estrellas4/5 (19)
- Investigación de Nuerologia Con APADocumento81 páginasInvestigación de Nuerologia Con APAEvelyn Edith Zetino Garc�aAún no hay calificaciones
- NEURULACIÓNDocumento4 páginasNEURULACIÓNAbraham SánchezAún no hay calificaciones
- NEURULACIONDocumento12 páginasNEURULACIONDulce Rayo LealAún no hay calificaciones
- Embriologia Del Sistema NerviosoDocumento3 páginasEmbriologia Del Sistema NerviosoCata MartinAún no hay calificaciones
- Tarea 1 de PsicofisiologiaDocumento16 páginasTarea 1 de Psicofisiologialaura michelleAún no hay calificaciones
- Sistema nervioso: Definición, estructura y desarrollo embriológicoDocumento15 páginasSistema nervioso: Definición, estructura y desarrollo embriológicoElexnat IdiomasAún no hay calificaciones
- Embriología Del Sistema NerviosoDocumento9 páginasEmbriología Del Sistema NerviosoBETSY ANDREA CHUQUIN GARAYAún no hay calificaciones
- Desarrollo embrionario de la tercera a la octava semanaDocumento12 páginasDesarrollo embrionario de la tercera a la octava semanaEnoc GonzálezAún no hay calificaciones
- Nervioso CKQDocumento33 páginasNervioso CKQMatty MarchandAún no hay calificaciones
- Neurulación EmbrioDocumento19 páginasNeurulación EmbrioCamila Calderón RioAún no hay calificaciones
- Neurulacion e Histogenesis Del SNDocumento7 páginasNeurulacion e Histogenesis Del SNJorge Luis Araujo III AAún no hay calificaciones
- Sistema Nervioso PDFDocumento18 páginasSistema Nervioso PDFJOCELYN FIERRO GUERREROAún no hay calificaciones
- Mecanismos de NeurulaciónDocumento7 páginasMecanismos de NeurulaciónGabriela VelásquezAún no hay calificaciones
- 12 Capas GerminativasDocumento22 páginas12 Capas Germinativasgiuseppeperoni PichardoAún no hay calificaciones
- MonofrafíaDocumento12 páginasMonofrafíaLucero Palomino RomeroAún no hay calificaciones
- Apuntes Embriologia Del Sistema Nervioso PDFDocumento7 páginasApuntes Embriologia Del Sistema Nervioso PDFIsi ·ᴥ·Aún no hay calificaciones
- Desarrollo de Sistema NerviosoDocumento8 páginasDesarrollo de Sistema NerviosoSebastián Andrés Urrea GonzalezAún no hay calificaciones
- Apuntes Embriologia Del Sistema NerviosoDocumento15 páginasApuntes Embriologia Del Sistema NerviosoCatalina Soler Lioi100% (1)
- Guía de Embriología Del Sistema NerviosoDocumento23 páginasGuía de Embriología Del Sistema Nerviosoprotoprueba123Aún no hay calificaciones
- Periodo Organogenético y FetalDocumento12 páginasPeriodo Organogenético y FetalAngie Lucia Llerena TorresAún no hay calificaciones
- Tercera semana de desarrollo embrionario: Gastrulación y formación de las capas germinativasDocumento42 páginasTercera semana de desarrollo embrionario: Gastrulación y formación de las capas germinativasjuanAún no hay calificaciones
- Catedra de Laboratorio de Neuroanatomía Embriologia e HistogenesisDocumento8 páginasCatedra de Laboratorio de Neuroanatomía Embriologia e HistogenesisGregory DislaAún no hay calificaciones
- 10 Segundo SISTEMA NERVIOSO 1Documento61 páginas10 Segundo SISTEMA NERVIOSO 1elpinchosblogAún no hay calificaciones
- Embriologia Del SNC - Filogenia y OntogeniaDocumento19 páginasEmbriologia Del SNC - Filogenia y OntogeniaSlim InfinitoAún no hay calificaciones
- Sistema nervioso centralDocumento7 páginasSistema nervioso centralGreysmar Vanessa Chirinos GuapeAún no hay calificaciones
- Desarrollo SNC y órganos sentidosDocumento5 páginasDesarrollo SNC y órganos sentidosmv glAún no hay calificaciones
- Desarrollo Embrionario y Neurobiologia de La Neurona y NeurogliaDocumento6 páginasDesarrollo Embrionario y Neurobiologia de La Neurona y NeurogliaLisbeth Alejandra Martínez GonzálezAún no hay calificaciones
- Desarrollo Embrionario SISTEMA NERVIOSO-1Documento70 páginasDesarrollo Embrionario SISTEMA NERVIOSO-1Brenda Gress ViverosAún no hay calificaciones
- Desarrollo SNC 38Documento3 páginasDesarrollo SNC 38AnaRamirezAún no hay calificaciones
- Guia 5Documento4 páginasGuia 5Ana Belén MedinaAún no hay calificaciones
- Resumen NeuroDocumento91 páginasResumen NeuroNa ParritaAún no hay calificaciones
- Sistema nervioso central 2.0Documento5 páginasSistema nervioso central 2.0Jorge Chz subsAún no hay calificaciones
- Lectura NeurodesarrolloDocumento10 páginasLectura Neurodesarrollozodiac_38Aún no hay calificaciones
- Resumen Embrio 3 Sistema NerviosoDocumento7 páginasResumen Embrio 3 Sistema Nerviosolucasreal.tapiaAún no hay calificaciones
- 06 - Periodo EmbrionarioDocumento49 páginas06 - Periodo Embrionariolovers of everytingAún no hay calificaciones
- Psicobiología Segundo Parcial (I)Documento112 páginasPsicobiología Segundo Parcial (I)Laura MurAún no hay calificaciones
- Desarrollo del Sistema Nervioso (SNDocumento20 páginasDesarrollo del Sistema Nervioso (SNcgilgas100% (1)
- Desarrollo del Sistema Nervioso CentralDocumento8 páginasDesarrollo del Sistema Nervioso CentralDiego Bedón AscurraAún no hay calificaciones
- Neurodesarrollo PDFDocumento18 páginasNeurodesarrollo PDFLissCarrilloAún no hay calificaciones
- Embriologia Del SNDocumento12 páginasEmbriologia Del SNAlondra SiguasAún no hay calificaciones
- Desarrollo Del Sistema Nervioso CentralDocumento3 páginasDesarrollo Del Sistema Nervioso CentralYuliana MendozaAún no hay calificaciones
- Embriología Del SNCDocumento5 páginasEmbriología Del SNCManuel Alejandro Parada100% (1)
- DE 3ERA A 8VA SEMANA DEL DESARROLLO - PERÍODO EMBRIONARIO (Embriologia Langmann 8ed)Documento6 páginasDE 3ERA A 8VA SEMANA DEL DESARROLLO - PERÍODO EMBRIONARIO (Embriologia Langmann 8ed)Maria Silva DiazAún no hay calificaciones
- Medula Espinal: Capa Neuroepitelial, Capa Del Manto y Capa MarginalDocumento12 páginasMedula Espinal: Capa Neuroepitelial, Capa Del Manto y Capa MarginalArturo TenorioAún no hay calificaciones
- Transcripcion Desarrollo Del Sistema NerviosoDocumento9 páginasTranscripcion Desarrollo Del Sistema NerviosoValentina EcAún no hay calificaciones
- Gould, D. (2015) - Neuroanatomía (5th Edition Ed.) y Purves de NeurocienciaDocumento5 páginasGould, D. (2015) - Neuroanatomía (5th Edition Ed.) y Purves de NeurocienciaFelipe GuzmanAún no hay calificaciones
- Embriología Del Sistema NerviosoDocumento12 páginasEmbriología Del Sistema NerviosoJuliana CarrilloAún no hay calificaciones
- Embriología Del Sistema NerviosoDocumento104 páginasEmbriología Del Sistema NerviosoDaniel Sebastian100% (2)
- TEMA 9Documento14 páginasTEMA 9arancharr01Aún no hay calificaciones
- Sistema NerviosoDocumento13 páginasSistema NerviosoGarzain ReyesAún no hay calificaciones
- Sistema Nervioso Central EmbrioDocumento16 páginasSistema Nervioso Central EmbrioRicardo Molina OrtizAún no hay calificaciones
- Embriologia NEURO ApuntesDocumento25 páginasEmbriologia NEURO ApuntesPepa PigAún no hay calificaciones
- Periodo EmbrionarioDocumento16 páginasPeriodo Embrionarioffloresp100% (6)
- HDocumento10 páginasHGeorge BetancourthAún no hay calificaciones
- De La Tercera A La Octava Semana Embrionaria y Hematopoyesis Fetal 327600 Downloable 519173 PDFDocumento19 páginasDe La Tercera A La Octava Semana Embrionaria y Hematopoyesis Fetal 327600 Downloable 519173 PDFJeffrey LoayzaAún no hay calificaciones
- Embriologia. Sistema NerviosoDocumento11 páginasEmbriologia. Sistema Nerviosowhitney63Aún no hay calificaciones
- Embriologia Del Sistema Nervioso CentralDocumento24 páginasEmbriologia Del Sistema Nervioso CentralGELEXSAún no hay calificaciones
- Anatomia y Fisiología del Sistema Nervioso IIDe EverandAnatomia y Fisiología del Sistema Nervioso IICalificación: 4 de 5 estrellas4/5 (28)
- Músculos de La Cabeza y El CuelloDocumento40 páginasMúsculos de La Cabeza y El CuelloJimena GuerrerosAún no hay calificaciones
- Maternizacion de LecheDocumento4 páginasMaternizacion de LecheJimena GuerrerosAún no hay calificaciones
- Semiología Cardiovascular y SoplosDocumento27 páginasSemiología Cardiovascular y SoplosJimena GuerrerosAún no hay calificaciones
- Articulaciones de Cabeza y CuelloDocumento33 páginasArticulaciones de Cabeza y CuelloJimena GuerrerosAún no hay calificaciones
- Osteología CaraDocumento34 páginasOsteología CaraJimena GuerrerosAún no hay calificaciones
- 6) Embriologia SISTEMA RESPIRATORIO-1Documento5 páginas6) Embriologia SISTEMA RESPIRATORIO-1Jose Guillermo Mejia SaldanaAún no hay calificaciones
- FIL ADD: Mecanismos Biológicos Del DesarrolloDocumento4 páginasFIL ADD: Mecanismos Biológicos Del DesarrolloJimena GuerrerosAún no hay calificaciones
- Nervios de la cabeza y el cuello: guía básicaDocumento38 páginasNervios de la cabeza y el cuello: guía básicaJimena GuerrerosAún no hay calificaciones
- Embriologia 3 EraDocumento2 páginasEmbriologia 3 EraJimena GuerrerosAún no hay calificaciones
- UROGENITALDocumento26 páginasUROGENITALJimena GuerrerosAún no hay calificaciones
- Cara Cuello Ojo y OidoDocumento27 páginasCara Cuello Ojo y OidoJimena GuerrerosAún no hay calificaciones
- 004 Sesión 004 Los 4 Temperamentos Fortalezas y Debilidades 1807Documento12 páginas004 Sesión 004 Los 4 Temperamentos Fortalezas y Debilidades 1807OTTO IVAN RODRIGUEZ RODASAún no hay calificaciones
- 10 Herencia y EvoluciónDocumento18 páginas10 Herencia y EvoluciónLisbeth Velasco RodriguezAún no hay calificaciones
- Triptico Celula BiologiaDocumento2 páginasTriptico Celula BiologiaImpresiones Trabajos EscolaresAún no hay calificaciones
- 10 Teorías Del Origen de La VidaDocumento7 páginas10 Teorías Del Origen de La VidaDanny Collado RodAún no hay calificaciones
- Guía de Estudio Biología L U 3 S 4Documento14 páginasGuía de Estudio Biología L U 3 S 4Jessica AguirreAún no hay calificaciones
- Taller HongosDocumento3 páginasTaller HongosAndres RodriguezAún no hay calificaciones
- Ácidos NucléicosDocumento9 páginasÁcidos NucléicosCyber El CoyAún no hay calificaciones
- Tarea 2Documento8 páginasTarea 2Peri-Hala Norbibi Monago SantosAún no hay calificaciones
- Clase 6 OpDocumento5 páginasClase 6 OpLucila HuamánAún no hay calificaciones
- OCR Lehninger Principios de Bioquimica 7a EdicionDocumento1235 páginasOCR Lehninger Principios de Bioquimica 7a EdicionFatima Torres100% (1)
- Precursores de La CriminologiaDocumento85 páginasPrecursores de La CriminologiaSergio Ochoa50% (2)
- Laboratorio NCBIDocumento20 páginasLaboratorio NCBIXIME GRANIFOAún no hay calificaciones
- Cómo Corregir La Salinidad Del SueloDocumento14 páginasCómo Corregir La Salinidad Del SueloAnays DíazAún no hay calificaciones
- Morfofisiologia Cintura Escapular y PélvicaDocumento13 páginasMorfofisiologia Cintura Escapular y PélvicaMelina ParragaAún no hay calificaciones
- CARTAS Y COMBINACIONES ESPECÍFICASDocumento5 páginasCARTAS Y COMBINACIONES ESPECÍFICASIván E. V.Aún no hay calificaciones
- 8-Texto Del Artículo-36-2-10-20191010Documento8 páginas8-Texto Del Artículo-36-2-10-20191010Alexander AlayAún no hay calificaciones
- Charolais-Juan MachadoDocumento12 páginasCharolais-Juan MachadosranAún no hay calificaciones
- Anatomía Del Hueso Del Húmero Del Perro AnatomyLearner - El Lugar para Aprender Anatomía Veterinaria en LíneaDocumento36 páginasAnatomía Del Hueso Del Húmero Del Perro AnatomyLearner - El Lugar para Aprender Anatomía Veterinaria en Líneaclips and chipsAún no hay calificaciones
- Cs. Naturales 5° Básico Guía N°15 Evaluación Sistema Respiratorio y CircularDocumento4 páginasCs. Naturales 5° Básico Guía N°15 Evaluación Sistema Respiratorio y CircularCynthia AvgAún no hay calificaciones
- Planclase - MATEMÁTICAS, - CUERPO - HUMANO - Y - BIODIVERSIDAD - Semana 4 (4 Marzo - 8 Marzo)Documento11 páginasPlanclase - MATEMÁTICAS, - CUERPO - HUMANO - Y - BIODIVERSIDAD - Semana 4 (4 Marzo - 8 Marzo)yeseniaAún no hay calificaciones
- Variaciones Linguisticas para Cuarto de SecundariaDocumento5 páginasVariaciones Linguisticas para Cuarto de SecundariaKL GTAún no hay calificaciones
- RCA-TCA05242-2022 - RCA-TCA05244-2022 - UnlockedDocumento3 páginasRCA-TCA05242-2022 - RCA-TCA05244-2022 - UnlockedSergio Amado EspinosaAún no hay calificaciones
- ZOO GITA UNI 10 Parte 1Documento14 páginasZOO GITA UNI 10 Parte 1joel rico aroniAún no hay calificaciones
- Laboratorios en ColombiaDocumento18 páginasLaboratorios en ColombiaJhosua MckenleyAún no hay calificaciones
- Ezequiel Ander EggDocumento26 páginasEzequiel Ander EggLuis BisetAún no hay calificaciones
- Resumen Yoga de Las VocalesDocumento7 páginasResumen Yoga de Las VocalesShanti GonzálezAún no hay calificaciones
- Braidot - Neurobiologia y Neurociencia en La Toma de DecisionesDocumento7 páginasBraidot - Neurobiologia y Neurociencia en La Toma de DecisionesAlyson CruzAún no hay calificaciones
- 2 - Guía Didáctica - C. Naturales - Sexto - MayoDocumento24 páginas2 - Guía Didáctica - C. Naturales - Sexto - MayoWill JJAún no hay calificaciones
- Guía 3Documento3 páginasGuía 3VictorAún no hay calificaciones
- 17-Dblm2bia4n3db0dhtbg9wtocs 20211109060442Documento3 páginas17-Dblm2bia4n3db0dhtbg9wtocs 20211109060442Adriana Estefania Tomalá GómezAún no hay calificaciones